 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 28 NUMBER 1 JANUARY 1989
Articles and Notes
Sustained Primate Conservation, by W. R. Kingston ...... 1
Preliminary Report on the Use of a Natural Foraging Task to Reduce Aggression and Stereotypies in Socially Housed Pigtail Macaques, by Maria L. Boccia ...... 3
Nylon Balls Re-visited, by Kathryn A. L. Bayne 5
Polyvinyl Chloride Piping as Perch Material for Squirrel Monkeys, by Axel V. Wolff 7
Rhesus Monkey Colony Management: A 1988 Update 8
Guidelines to Prevent Simian Immunodeficiency Virus Infection in Laboratory Workers and Animal Handlers 17
Male Rhesus Monkey's Unusual Behavior: Interspecies Adoption, by Victor E. Collazo 23
News, Information, and Announcements
Fyssen Foundation 1989-1990 Fellowships and 1989 International Prize 2
Errata and Amplifications 4
Meeting Announcements 14
. . IPS, Fertility in Great Apes, NABR, PHS Policy Workshop, APS
Letters: Endangered Chimps 15
Information Available 16
. . AIDSLINE, Animal Welfare Information Center, PSIC
Research Reports 22
. . Pulmonary Hypertension, False-Positive AIDS Tests
News Briefs 24
. . Chimpanzee Sperm Bank, Lawsuit Against U. C. Dismissed,
Pickett Retires, ICLAS, NIH Briefs Congressional Aides,
Costs for New Animal Rules.
Cartoon ...... 23
Departments
Address Changes 25
Positions Available 26
Recent Books and Articles 27
* * *
W. R. Kingston
There has recently been considerable publicity for the "Debt for Conservation" schemes, under which heavily discounted foreign debt notes are bought up by sponsors in the United States, converted back to local currency, and the much enhanced proceeds donated to government agencies promising to use them for "the purchase and maintenance of reserves including the provisions of guard posts and guards." This is undoubtedly a very good idea, and the very generous response of many U.S. citizens both to this and to World Wildlife Fund appeals shames the U.K. and western Europe generally. However, having spent the major part of the past twelve years working on conservation-oriented primate projects in Peru and Brazil, I am convinced that care needs to be taken to utilize this money to the best advantage rather than seeing it frittered away with no lasting benefits, at least to nonhuman primates.
There is no doubt that conservation of the whole habitat rather than one particular species should be the objective. The vast majority of species of platyrrhine primates occur in at least one of the reserves listed in the IUCN Directory of Neotropical Protected Areas, and most of these are government owned. There should therefore be very little need to purchase land for the protection of any species, the one obvious exception being the case of Brachyteles in Brazil, the main population of which is on a privately owned estancia.
With regard to guards or wardens, the current procedure of paying some local man a pitifully low salary and providing neither suitable equipment, transport, nor back-up facilities is worse than useless. To be anything like effective, there must be clear delineation of the duties and area under his charge; he must be paid a wage well above the local average to make it obviously to his advantage to keep his job; guards should patrol in pairs who should not be related. There must be a proper H.Q. building manned by a supervisor with wireless communication to the individual guards, who should be required to check in from suitably spaced posts on their territories at frequent intervals. Suitable transport``4 wheel drive vehicles or powered river boats``must be provided and liaison with the nearest police or military post kept open at all times. All this may sound very expensive, but even a very good local salary is cheap by U.S. standards: $100 (U.S.) per month is much above the current levels. The transport and other equipment would cost a lot less than the purchase of thousands of hectares of land. The funding authority should have the right to visit any site unannounced at any time, and to receive properly kept accounts of expenditure.
In practice, I do not think the concept of guards will ever be effective in most reserves in Amazonia unless the cooperation and involvement of local populations as a whole can be achieved. This will only come about if it can be demonstrated to these relatively uneducated but very practical people that conservation of the area gives them easily appreciated greater material benefit than its destructive exploitation. They cannot be expected to appreciate the concern for some small inedible and unsaleable creature, however important it may be to some remote "gringo," or to realise that the very tasty Ateles or Lagothrix they have hunted all their lives cannot reproduce fast enough to keep pace with improved hunting results brought about by the increasing availability of cheap shotguns. Their cooperation will be achieved if the reserve can provide regular work, either directly or by the provision of raw materials, such that the whole community benefits. Given this, they will support the guards and oppose destructive practices by anyone, including their own government.
The other essential requirement for continuity is the generation, or at least provision, of a regular income which must not be dependent on continuous appeals to the public of the developed nations. As I have said in an earlier paper (Kingston, 1985), machinery and equipment wears out and staff must be paid every month. There is a limit to public response, particularly when human tragedies of famine, flooding, and earthquakes result in an almost continuous appeal for funds, and we are quite obviously failing to control the suicidal population increases which are the greatest threat of all. Surely selected relatively small areas of the richest ecosystem in the world can be so managed so as to earn their own keep in a nondestructive manner. I am quite certain that this can be achieved in certain existing reserves in both Brazil and Peru, and at a cost well within the figures being published as available. A more detailed account of my proposals will be published elsewhere.
Reference
Kingston, W. R. (1985). Conservation of the Callitrichidae. Laboratory Primate Newsletter, 24[1], 1-3.
-------------------------------------------------------------------
Author's address: The Old Smithy, Bishops Frome, Worcester WR6 5BA, England.
* * *
1989-1980 Fellowships
The Fyssen Foundation's general aim is "to encourage all forms of scientific enquiry into cognitive mechanisms, including thought and reasoning, underlying animal and human behavior, into their biological and cultural bases, and into their phylogenetic and ontogenetic development." For this purpose, the Foundation will award a number of fellowships. These fellowships are meant for the training and support of research scientists working in disciplines relevant to the aims of the Foundation, which wishes to support, particularly, research in such fields as: Ethology and Psychology: Nature and development of the cognitive processes in man and animals. Neurobiology: Neurobiological bases of cognitive processes and of their embryonic and post-natal development, as well as the elementary mechanisms which they involve. Anthropology-Ethnology: Study of a) the representations of the natural and cultural environment. Analysis of the construction principles and transfer mechanisms of these systems as they illuminate their cognitive aspects, b) the technological systems developed in the various forms of social organization. Human Paleontology: Origin and evolution of the human brain and human artifacts.
Fellowships will be given to French scientists to work abroad and to foreign scientists to work in French laboratories. Study grants will normally be granted for one year but may be extended up to three.
Application forms can be obtained from the Foundation. Applications must include: a curriculum vitae; a list of publications of the applicant; the names of two senior scientists whom the applicant has asked to send testimonials to the Secretariat of the Foundation by March 31, 1989; a letter of acceptance of the inviting laboratory. Fifteen copies of the completed information should be sent to the Secretariat of the Foundation, 194 rue de Rivoli, 75001 Paris, France. Deadline for receipt of applications by the Foundation is March 31, 1989.
International Prize
A substantial International Scientific Prize will be given for a major contribution to the progress of knowledge in the fields of research supported by the Foundation. It was awarded to Professors Andre Leroi-Gourhan in 1980, William H. Thorpe in 1981, Vernon B. Mountcastle in 1982, Harold C. Conklin in 1983, Roger W. Brown in 1984, P. Buser in 1985, David Pilbeam in 1986, and David Premack in 1987. The discipline considered for the 1989 prize is Neurosciences. The nominations should include a curriculum vitae of the nominee; a list of his publications; a summary (four pages maximum) of the research work upon which the nomination is based. 15 copies of the nomination should be sent to the Secretariat of the Foundation at the above address. Deadline for receipt of nominations is September 1, 1989.
* * *
Maria L. Boccia
University of Colorado Health Sciences Center
Introduction
Nearly all nonhuman primates are highly social. Among the most significant elements that can be introduced for enrichment, therefore, are social companions (O'Neill, 1988; Reinhardt, Eisele, & Houser, 1988). Provision of social companions, however, may introduce several potential hazards. In addition to the problem of disease transmission, the prevalence of dominance hierarchies introduces the potential for competition for food, water, and preferred perch sites, as well as aggression leading to injury. The study reported here examines the impact of providing a natural foraging task on the agonistic behavior of socially housed pigtail monkeys.
Methods
A group of socially housed pigtail monkeys (Macaca nemestrina) , consisting of an adult male, 7 adult females, and 7 juveniles and infants, was studied. Water was freely available and food was provided at 0900 hr by scattering monkey biscuits and fresh fruit across the floor of the pen. The pen was 2.1x2.5x4 m. with cinderblock walls, wire mesh ceiling, and woodchip bedding on the cement floor. Additional space was provided with shelving and pipes.
Foraging consisted of a supplementary feeding of approximately one cup of sunflower seeds in the middle of the afternoons, 4-6 hours after the group was fed their daily ration of chow and fruit. The seeds were dispersed throughout the cage in the woodchip bedding. Animals were thus permitted to forage, by searching through the bedding, to find the seeds. For 2 weeks before and after beginning this regimen, five-minute focal animal samples of behavior were obtained from 5 females 9-10 years old, 3 females 4-5 years old, and 3 juveniles 1-2 years old. These data, including social and exploratory behaviors, were subjected to ANOVAs with condition (pre/post introduction of seeds) and age as factors.
Results
With the introduction of the foraging task, social, exploratory, and abnormal behaviors changed. Stereotypies decreased (p = .04), and hairpulling, which prior to the introduction of the seeds occurred primarily in the afternoons, was nearly eliminated. In addition, bedding (p = .003) and other types of exploration (p = .05) increased, agonistic behaviors decreased (fight: p = .01; aggress: p = .08; submit: p = .01), and play increased (p = .01) (see Figure 1).
Figure 1: Behavioral responses of all monkeys observed in response to the introduction of sunflower seeds (Loc = locomote; AS = attempt to search outside pen; OE = explore objects; BE = explore bedding; SM = stereotypic motor behavior).
Several behaviors were differentially affected, depending on the age class of the monkeys: juveniles showed the greatest increase in play (p = .01) and decrease in locomotion (p = .008); adults the greatest increase in bedding exploration (p = .04). Affiliative behaviors showed different changes depending on the monkey's age: juveniles showed a decrease, while the adults and subadults showed an increase (p = .04) (see Figure 2).
Figure 2: Different responses of adults, subadults, and juveniles to introduction of sunflower seeds.
Discussion
Feeding strategies may compound or reduce the hazards of housing primates in social groups. Given the trend toward social housing as an enrichment strategy for laboratory primates, it is imperative to discover techniques which can be easily and inexpensively implemented ,cb and which will minimize aggressive behavior and the attendant risk of physical injury. The presence of a food which requires a natural foraging activity not only reduced such agonistic behavior and increased play, but also reduced stereotypies, including hairpulling behavior in the pigtail macaques studied here. The effects of this manipulation were somewhat affected by the age of the monkeys, suggesting that the impact of enrichment strategies will have to be assessed for differences across age and possibly sex, as well as for species differences.
It appears to be possible, however, to use social housing as a means of enrichment for primates, and incorporate management procedures that will reduce the levels of stereotypies, and the potential hazards of agonistic interactions. A follow-up study is currently in progress to determine if this foraging task continues to be effective in reducing aggression, hairpulling, and stereotypies on a longterm basis.
References ;
O'Neill, P. (1988). Developing effective social and environment enrichment strategies for macaques in captive groups. Lab Animal, 17[4], 23-36.
Reinhardt, V., Eisele, S., & Houser, D. (1988). Environmental enrichment program for caged macaques: A review. Laboratory Primate Newsletter, 27[2], 5-6.
-------------------------------------------------------------------
Author's address: Department of Psychiatry, C-268R,
University of Colorado Health Sciences Center, 4200 East Ninth Ave.,
Denver, CO 80262.
This research supported by USPHS grant MH 44131.
* * *
Environmental Enrichment
In the announcement "Environmental Enrichment Slides and Videotape," on page 4 of the last issue, the phone number for the WRPRC Library Visual Services was given incorrectly. The correct number is 608-263-3512.
Maternal Behavior
Bernadette, the chimpanzee described in the article "Competent Maternal Behavior by a Hand-Reared, Resocialized, Primiparous Chimpanzee: A Case History," by Nankivell et al., on pages 5 to 8 of the last issue, was born and spent the first 31 months of her life at the Laboratory for Experimental Medicine and Surgery in Primates (LEMSIP), New York University School of Medicine. Rearing of newborn infants at LEMSIP is described in "Chimpanzees in Captivity: Humane Handling and Breeding within the Confines Imposed by Medical Research and Testing," J. Moor-Jankowski and C. J. Mahoney. Journal of Medical Primatology, in press.
* * *
Kathryn A. L. Bayne
National Institutes of Health
Renquist and Judge (1985) have recommended placement of nylon balls&S'1. in the cages of singly-housed nonhuman primates for the purpose of environmental enrichment and prevention or reduction of abberant behaviors. The absence of any specific description by Renquist and Judge of the ways in which nonhuman primates use nylon balls prompted this author to further investigate the kinds of interactions the animals have with this device. In a comment appended to Renquist and Judge's article, the editor of this journal indicated that from a management standpoint the potential for noise disturbance might be a negative outcome of the incorporation of this enrichment device in nonhuman primate cages. Renquist and Judge also noted one instance of a ball that blocked a floor drain, and the destruction of a $7000 sewer pump has been reported to this author. These illustrations aptly point out that no enrichment device is without its drawbacks, some of which can have unexpected economic impacts.
Informal observations of the nonhuman primates in two animal rooms were made for 30 to 90 minutes between 1000 and 1600 hrs, two to three times a week over a period of three months. The animals in the rooms had been maintained in the facility from less than one year (for infant and juvenile monkeys) to five years. They ranged in age from less than one year to 27 years. As the animals came from a variety of sources (both feral and captive born), they represented a broad scope of rearing conditions (nursery, nuclear family, peer, mother/infant, mother/peer). Each cage in this particular facility at the NIH is routinely equipped with a nylon ball; therefore all animals had been exposed to the ball from the time they were placed in their cages. It may be assumed, then, that any consistent interaction between an animal and the ball was not due to a novelty effect.
* Animal sits on ball, occasionally with legs propped up on cage wall * Animal holds ball with hands or feet * Animal chews on/bites ball * Animal holds ball in mouth while sitting or moving around the cage * Animal picks at seam of ball * Animal paces around cage and moves ball with hand with each pass by it * While sitting, animal repeatedly rolls ball away and watches it roll back * Animal rubs ball on cage floor, thereby generating noise * Animal repeatedly picks up and drops ball
Table 1: Observed interactions between nonhuman primates and nylon balls.
During each half-hour period a minimum of three animals out of an average of 34 animals in a room (different animals during different observation times) manipulated the nylon ball. This figure of 10% of the rhesus population coincides with what Line (1987) has reported. During a single half-hour observation period seven animals (19% of the animals in the room) interacted with a ball. The various ways in which the rhesus (Macaca mulatta) , stumptail (M. arctoides) , and cynomolgus monkeys (M. fascicularis) used the nylon ball are described in Table 1. As can be expected with any enrichment tool, some individual animals routinely manipulated the nylon balls while others interacted with it less frequently or not at all. Occasionally the interaction was part of a stereotypic pattern (e.g., circling around the cage and touching the ball with each pass), so it may be suggested that for those particular animals the presence of the nylon ball has not eliminated all the abnormal behavior patterns manifested by these animals. Also, one animal consistently incorporated the nylon ball into a self-abusive behavior pattern, alternately biting his leg and the nylon ball.
An environmental enrichment device like the nylon ball helps elucidate features of devices in general which are important to the animal. Since the design of the nylon ball is very simple, it may be easier to identify those features of it which are the most salient and interesting (or "species appropriate," as Line refers to it) to the animals. These features may then prove to be important in directing the design of future enrichment devices aimed at improving the psychological well-being of nonhuman primates.
The concept of improving psychological well-being by providing an animal with control over its environment is often referred to by investigators in this field (Novak & Drewsen, 1988; Line, 1987; Mineka et al, 1986; Curtis, 1985). Does the nylon ball confer to the nonhuman primate some control over the cage environment (e.g., by using it to generate noise)? If it is regarded by the animal as an extension of the cage, then it is one of the few elements that is moveable. This mobility or portability may be an attractive feature of the ball. Baldwin and Suomi (1974) have found that infant rhesus monkeys explored moveable objects more frequently and for longer durations than stationary stimuli. For example, many animals, when given the opportunity to do so, will operate the squeeze apparatus of their cages by themselves on a regular basis. This, too, may be a demonstration of the animals' need to be able to alter the environment themselves, or live in an environment which is changeable to some degree. Often the first reaction of a nonhuman primate to its new cage is to pick up the nylon ball and cling to it while moving around the cage (M. April, personal communication).
Even if the nylon ball is not regarded by the animals as an extension of the home cage per se, it is certainly viewed by some animals as "property" or part of the home territory. Animal care and laboratory technicians have reported to this author how difficult it can be to remove a nylon ball from an animal's cage, as the animal will often grab the ball and hold it away from the individual attempting to remove it. Do enrichment strategies that require the rotation of devices or "toys," then, inject an element of stress into a program designed to improve "psychological well-being" by periodically removing part of an animal's territory and substituting a novel object?
Since approximately 10% of the monkeys in a room utilize the ball at any given time, the consistency in appeal of the nylon ball is not equal for all macaque species or individuals. Yet, many animals interact with the ball on a regular basis. As can be expected with any enrichment device, then, the nylon ball is not a ubiquitous solution for enhancing the psychological well-being of nonhuman primates. For researchers, the nylon ball may be useful as a tool to assist in the identification of those features of a device which are interesting to nonhuman primates and which could be incorporated in novel enrichment devices and strategies.
References
Baldwin, D. & Suomi, S. (1974). Reactions of infant monkeys to social and nonsocial stimuli. Folia Primatologia, 22, 307-314.
Curtis, S. (1985). What constitutes animal well-being? In G. A. Moberg (Ed.), Animal Stress (pp. 29-49). American Physiological Society.
Line, S. (1987). Environmental enrichment for laboratory primates. Journal of the American Veterinary Medical Association, 190, 854-859.
Mineka, S., Gunnar, M., & Champoux, M. (1986). Control and early socioemotional development: Infant rhesus monkeys reared in controllable versus noncontrollable environments. Child Development, 57, 1241-1256.
Novak, M. & Drewsen, K. (1988). Enriching the lives of captive primates: Issues and problems. In E. Segal (Ed.), Psychological Well-Being of Captive Primates. Noyes (In press).
Renquist, D. M. & Judge, F. J. (1985). Use of nylon balls as behavioral modifier for caged primates. Laboratory Primate Newsletter, 24[4], 4.
-------------------------------------------------------------------
Author's address: Department of Health & Human Services,
NIH, Building 14D, Room 309, Bethesda, MD 20892.
®Nylaball, Nylabone Corp., P.O. Box 27, Neptune, NJ 07753.
Note that unlike what Line (1987) has reported, in
these recordings cynomolgus monkeys were observed using the nylon ball.
* * *
Axel V. Wolff
National Institutes of Health
Environmental enrichment and methods to encourage and facilitate primates to exhibit species-specific behavior are issues receiving increasing attention (Reinhardt & Smith, 1988; Reinhardt et al, 1987; Line, 1987). Polyvinyl chloride (PVC) piping has been successfully used for primate cage enrichment by various institutions. The present article illustrates how our facility utilizes this material as perches for squirrel monkeys (Saimiri sciureus).
A colony of 250 squirrel monkeys of both sexes and of varying ages was studied. They are at the Laboratory of Central Nervous System Studies, National Institute of Neurological and Communicative Disorders and Stroke, animal facility in Frederick, MD. The animals were housed one to four in each standard stainless steel (Harford Metal Products, Inc.) primate cage measuring 20x22x34 in.
To enrich the cage environment, PVC piping was installed. The 1/2-inch internal diameter piping, cut to lengths of 20-1/4 inches, was capped at both ends. The caps allowed the piping to be wedged securely into the cage, but facilitated rapid removal without the use of tools. The caps also effectively sealed the interior of the pipes. The small size of the squirrel monkeys prevented them from moving or dislodging the perch in any way. The perch became a part of the cage and could be easily cleaned in place. However, since disinfection in 180deg F water in an automatic cage washer tends to distort the shape of the perch, removal of the bar and soaking in a mild sodium hypochlorite solution was preferable.
Initial reaction to the perch on the first few days was one of apprehension, followed by curiosity, and finally acceptance and usage. Within one month of introduction, usage of the perches was exhibited by all animals at some time during the daily routine. Perch usage consists of sitting, eating, sleeping, and locomotion in a fashion similar to tree limb use in wild monkeys. Occupation of the perch can be by one or all cagemates. Aggression for possession or control of the perch has not been noted.
Photo: Squirrel monkey on PVC perch eating egg.
Sanitary perches for small arboreal primates, such as squirrel monkeys or marmosets, encourage species-specific behavior without compromising routine husbandry or sanitation procedures. Cost, installation, and maintenance are minimal. Larger species need more permanently anchored perches. An additional unexpected benefit of the PVC piping has been a decrease in dorsal tail-head abrasions, frequently seen in squirrel monkeys that sit on the stainless steel flooring of standard primate cages.
References ;
Reinhardt, V. & Smith M. (1988). PVC pipes effectively enrich the environment of caged rhesus monkeys. Laboratory Primate Newsletter, 27[3], 4-5.
Reinhardt, V., Houser, W. D., Cowley, D., & Champoux, M. (1987). Preliminary comments on environmental enrichment with branches for individually caged rhesus monkeys. Laboratory Primate Newsletter, 26[1], 1-3.
Line, S. W. (1987). Environmental enrichment for laboratory primates. Journal of the American Veterinary Medical Association, 190, 854-859.
-------------------------------------------------------------------
Author's address: Laboratory of Central Nervous System Studies, NINCDS, National Institutes of Health, Bethesda, MD 20892.
* * *
Introduction
This workshop is a follow-up to a symposium on colony management of rhesus monkeys that I organized for the 1986 ASP meetings in Austin, TX. We had representatives from five of the NIH supported Primate Centers (Oregon, Washington, California, Yerkes, Delta), Yemassee, and the Caribbean Primate Research Center. At that time, it was strongly suggested that we continue the exchange of information, hence, the present workshop. This year we see the addition of the New England Primate Center, as well as representatives from two commercial breeders of macaques.--Margaret R. Clarke.
Charles Rivers Key Lois
I'll give you a brief overview of the Charles Rivers Colony in the Florida Keys. Key Lois was started in 1973 as a commercial venture, and in 1976 we started a colony with the FDA on Raccoon Key. Our base of operations is Summerland Key, about 25 miles north of Key West. Key Lois is about 100 acres, about 3 km out on the Atlantic side, and has about 1500 rhesus. Raccoon Key is about 200 acres, about 5 km out on the Gulf side, and has about 2200 rhesus.
The main problem we're having right now is defoliation. The islands have become overpopulated, especially Key Lois, which has really been stripped. We are reaching agreements with the Departments of Environmental Regulations and of Natural Resources of the state of Florida, whereby we will reduce the free-ranging population by putting a few corrals on the island. I hate to get into that, but there is no alternative if we are to meet our commercial obligations. So we will, in the future, have a combined corral/island operation.
The other problem with our colony is that we should have had more planning for replacement breeders. A lot of our native Indian animals are reaching 17-18 years of age and the birth rate is starting to drop. We are going to have to concentrate on replacing breeder stock. Along with the corral operation, we're not sure exactly what that is going to do to us, but I'm not taking any new orders right now.--Paul Schilling, Charles Rivers Key Lois, Box 259, Summerland Key, FL 33042.
Yerkes Regional Primate Research Center
As we reported at the Austin meetings in 1986, a series of overthrows of the highest ranking matriline in one of our rhesus groups took place between January of 1976 and February of 1983. Three of these 4 overthrows occurred between late 1982 and early 1983. These were the result of predominantly adult female members of the multiple lower ranking matrilines jointly attacking the adult female and adolescent members of the then-highest-ranking matriline of the group. In the 3 overthrows of 1982 and 1983, the next-ranking matriline assumed highest position, while the surviving members of the deposed matrilines fell to, or near, the bottom of the hierarchy. The group has been under continuous study since the last overthrow in 1983 and, in sharp contrast to the instability of late 1982 and early 1983, we can report that there have been no similar matrilineal overthrows, nor any significant changes in the matrilineal rankings since that time. Female dominance relationships have remained quite stable for the last 5 years, aside from expected maturational changes, and these also have been largely predictable in terms of youngest sister ascendancy.
We also reported in 1986 that the lack of participation in the overthrows by adult males, and their ineffective defense of the members of the attacked alpha matriline demonstrated independent functioning of the female core of the group from these adult males. Further evidence of this was noted in the fall of 1985, when all of the adult male members of the group were removed as part of an on-going research project. Their removal did not lead to any reorganization of female dominance relationships and the relative rankings of the matrilines have remained stable.
The relative positions at the bottom of the matrilineal hierarchy of the members of the overthrown matrilines which have remained in the group have also remained fairly stable, with one notable exception. A female of the alpha matriline overthrown in late 1982 (who was orphaned at that time when she was less than a year old) is now fully mature and has risen in rank to a position within that of the alpha matriline. This was a consequence of a relationship that she formed with an adult female member of the alpha matriline and despite resistance on the part of several mid-ranking adult females. After obtaining her high rank, she then aided her older sister, who began challenging higher ranking females, and she, too, rose drastically in rank. There is nothing to suggest, however, that other female members of the deposed matrilines will attain positions held by their mothers at the time of the overthrows. This is true for females who were adults when their matrilines were overthrown, as well as for those who were infants or very young juveniles at the time.
I think these observations recorded in the years since the series of overthrows support three basic conclusions regarding this group: First, this group is in no way characterized by what might be labeled "long term instability or abnormality." Second, young animals of overthrown matrilines can remain viable members of their group after the violent depositions of their matrilines, although they are likely to remain low ranking. Finally, not only were the violent reorganizatons which took place in the past independent of the adult males, but the stability which followed the overthrows has been maintained for over 5 years now, despite removal of all of the adult male group members in 1985.--Carolyn Ehardt, Department of Anthropology and Linguistics, University of Georgia, Athens, GA 30602.
I'd like to share a little anecdote with you. When we removed all of the adult males in 1985, the only males left in the group were immature natal males. Our alpha female's younger brother became the alpha male, but was, and still is, subordinate to his sister, although he was dominant to all of the males in the group. At age 4-1/2, he appeared to be the absolute alpha male in that group, and everything seemed to be perfectly in order until, one day, to our total surprise, a bunch of males in the group ganged up on him, attacked him, and beat him quite severely. We pulled him out immediately. He had minor injuries, but some of the cuts were deep. His matriline had come to his aid, and several of them were roughed up a bit, but his matriline retained status. When we returned him to the group he was immediately the target of attack by all of the other natal males. His matriline came to his aid at once. His sister sat directly in front of him and just stared at any male that tried to get to him, which turned out to be very effective. His aunt, a much larger and more vigorous female, physically attacked any male who came near him. Since she was about 7 kg and was attacking 8 to 9 kg males, she got roughed up, and then couldn't defend her nephew against other attacking males. So her vigorous attack was far less effective than the passive defense his sister provided.
We also noted something that had been reported previously. The female members in the matriline defended the males, while the alpha female's 2-year-old nephew and a very vigorous 4-year-old cousin led all the other males past their aunt and their mother to attack the alpha male. It appeared to us that other females were beginning to take advantage of that by moving in, and we feared that the alpha matriline might be overthrown in defense of this male. At this point, we elected to pull him out of the group. That was 1-1/2 years ago, and the alpha matriline has re-established itself; everything is as it was before. We felt that the male attack on this male did jeopardize the stability of the alpha matriline, and his removal re-established stability in the group. This demonstrates once again that female members are faithful to the matriline, whereas males are not. They are independent and fight quite freely with each other regardless of matrilineal affiliation. The matriline may give some support to a male, but the dominance position of a matriline does not seem dependent on the support or dominance of its male members.--Irwin Bernstein, Department of Psychology, University of Georgia, Athens, GA 30602.
New England Primate Research Center
The timed-mating program at the New England Regional Primate Research Center (NERPRC) includes 170 rhesus females and 25 males. We assign 60 of their offspring each year for fetal and developmental research, returning another 30 to 40 to our colony. This program is now producing its fourth generation.
NERPRC is reorganizing to allow more social contact among the program's females. Construction is nearly complete on a new building giving animals living in 15-18 m&S'2 rooms access to outdoors. Each room will house 3-5 females. Individuals move to a breeding room around the time of ovulation. We anticipate two main areas of difficulty with this program: a) the transition from the single cage environment to the small group setting; b) regular changes in the composition of the groups as individuals move to and from breeding rooms.
In 1987 we established a primate ethology unit to carry out ongoing studies of behavior of the monkeys to establish protocols for reintroductions and group formation for long term individually housed monkeys. This works as 4 components: 1) retrospective searches of the animal records to establish the extent and nature of their social experience; 2) applying one or more behavioral assessment scales; 3) observing their interactions in groups of 2 or 3; and 4) comparing the success of each individual with data collected in the other steps to develop a predictive model for successful resocialization.
We followed these steps in the establishment of a specific-pathogen-free colony in late 1987, consisting initially of about 25 rhesus monkeys with low or absent titres to type D retrovirus and Herpes B virus. This preliminary test of the protocol for social group formation from individually-housed animals was successful. To date, we've removed 1 female who was overly aggressive, 2 who never integrated into the group, and 1 whose B virus titre increased. Last year this colony produced 3 births, and we have added about 60 more juvenile rhesus and crab-eating monkeys in 3 other groups.
We are especially interested in the experiences of others who have made the transition from single to group-housed animals on a similar scale. We would also like to hear from colleagues using a variety of behavioral assessment methods to plan and monitor the progress of these moves.--Andrew J. Petto and Lyna M. Watson, NERPRC, One Pinehill Drive, Southborough, MA 01772.
Oregon Regional Primate Research Center
The first rhesus breeding corral at the Oregon Regional Primate Research Center was constructed in 1974. This 1-acre corral was initially stocked with 55 adult females and 13 adult males, which were a mix of wild-caught animals from India and colony-reared animals from the Center. In the following 3 years, 5 additional corrals were constructed. Maintenance of the corrals continued uneventfully until 1981, when a significant amount of fighting occurred among females. Genealogical analysis of animals involved in episodic aggression suggested matrilineal overthrows.
A number of different procedures were initiated in an attempt to reduce the fighting: 1) Grain was scattered to distract the animals. 2) Barrels were placed in the corrals to serve as hiding places. 3) The policy of treating and then returning injured monkeys was modified. If a monkey was removed for more than a week, it was not returned. 4) The number of adult males was increased in the corrals to control the fighting. 5) Animals were harvested from low-ranking matrilines, rather than equally from all matrilines.
Despite these changes in management practices, bouts of serious aggression continued. Two years ago a new management policy was instituted that involved eliminating matrilines. Two corrals were stocked with 2- and 3-year-old unrelated females. Current plans are to harvest all the yearling males, and all the 2-year old females every year from each corral. After a period of 6 to 8 years, corrals will be decomissioned in rotation, and the remaining adult females will be transferred to the Center's timed breeding colony or to research projects. New corrals will again be formed with a mix of 2- and 3-year-old females that have been harvested from other corrals.
This program appears to be working well. Birthrates in the two newly constituted corrals have been high, and there have been no significant episodes of serious aggression. However, two problems have occurred. 1) There is a relatively high rate of infant mortality among primiparous females. Differences in maternal behavior have been reported between multiparous and primiparous rhesus females and these will be investigated. 2) The corrals were stocked with age-graded cohorts of males (i.e., several 2-year-olds, several 3-year-olds, etc). When the young males reached puberty they were attacked by older males. Current plans are to put fully adult males in a corral. These males will be left as a group after the corral has been decomissioned, and then they will be released into a new corral. Separate groups of adult males will be rotated among the corrals to reduce inbreeding.
This program is supported by RR-0163 from NIH, USPHS.--G. Gray Eaton, Michael Axthelm, Stephen T. Kelley, & W. P. McNulty, Oregon Regional Primate Research Center, Beaverton, OR 97006.
Delta Regional Primate Research Center
This is an update on introductions of adult males into corral-living groups of rhesus monkeys at the Delta Primate Center. The FDA rhesus monkey breeding colony, consisting of 22 one-half acre corrals, had been stocked in 1974 with 5 males and 45 females each (sex ratio 1:9). By 1985, the sex ratio in some was as high as 1 male to 35 females. In the Fall of 1985, we formed 5 all-male groups, each with 5 to 9 fully adult males. They were housed in catchpens adjacent to corrals needing males, and were given a period of time to adjust to each other. Females from the adjacent corral were introduced, a few at a time, for 1-week periods, until they had all been with the males, and then the males were released into the corral. This approach failed for two reasons; the males became competitive in the presence of females and started fighting, and the females were attacked by group members once they were returned to the corral. Four of 5 females in one group were dead within a few hours of being returned to their corral, despite the fact that they never had been out of visual contact. In conjunction with these two problems, this procedure did not result in any improvement in the number of males being successfully introduced into established social groups.
In the Fall of 1986 and 1987, with advice from Irwin Bernstein, we implemented a new program which has been more successful. We target corrals with no or few males for replacement males. Those males are removed one full month before the start of the breeding season, and are housed in groups of 4-6 in catchpen areas. Peer groups of maturing males (4-5 years of age) are also housed in catchpen areas. Males are introduced approximately 1 month after the females start cycling. Males are released into a group with which they have had no social experience, which in our case also means that they are not released into the corral adjacent to their catchpen. The timing of the introduction is crucial as it must coincide with the breeding season for females who have not been with males. The only time free-ranging rhesus females accept males into the social group is during the breeding season, and this behavior is unchanged in corral-living situations. The males are released early in the day into their target corrals, and they are observed until dusk. If there are problems, the males are returned overnight to the adjacent catchpen and are released again in the morning.
In the Fall of 1987, we introduced 52 males. Twenty-two of them (42%) were accepted and now are living in the corrals. Twenty-one were pulled out because it was obvious that they were not integrating into the group, and these are being housed in peer groups until October, 1988. Nine animals were euthanized for trauma injury, but none were killed outright by group members.
The success of introduction correlated with the origin of the all-male group. We had old colony males, maturing Delta-born males, maturing Hazleton males who had always lived in social groups, and maturing Davis males who had been born in social groups but had been singly housed since 1 year of age. All of the younger males had been housed in catchpens at Delta for 4-6 months prior to introductions. The group of 3 old males entered their new group without incident, and they continued living there throughout the breeding season. Of the 14 Delta-born males, only 3 were successful at group membership, and remained in the corrals. Four were euthanized for trauma, and 5 are waiting for another chance this next year. Of the Hazleton/Davis animals, there was a 46% success rate. Four of the Davis males are in the group, 4 are waiting for this next year. Twelve of the Hazleton males are still in social groups, 10 are waiting for next year, and 5 were euthanized. Thus, the older males were the most acceptable, but the next most acceptable were complete strangers. There were no significant differences between the socially-housed Hazleton males and socially-then-singly-housed Davis males.
We will have more data next Fall. Many old males are dying of old age and must be replaced. We are particularly interested in looking at the success rate for animals rejected last year compared to first time introductions, in hope of identifying characteristics of males not having the social or physical expertise to successfully join an established group. We will continue to develop a plan for the proper combination of timing, group composition, and individual characteristics to maximize success of male introductions.--Margaret R. Clarke and James L. Blanchard, Delta Regional Primate Research Center of Tulane University, Covington, LA 70433.
Hazleton--Texas Primate Center
The Texas Primate Center is located approximately 40 miles west of Corpus Christi, TX. It is a 208 acre facility with 688 corncrib type cages, housing approximately 6000 macaques; 750 are cynomolgus, the rest are rhesus. Breeding harems are comprised of 1 male to an average of 7 females. The facility is divided up into colony areas, with a colony support building provided for each area.
The first procedure to be discussed is the return of convalescing adults back into the colony. Because of our limited hospital space, it is imperative that we return animals back to the harems as quickly as possible. We select animals, particularly adult females that have completed medical treatment but are still in need of some convalescence, for placement in corn cribs with 10-12 juvenile weanlings. One to 3 females will stay in that crib until fully recovered and strong enough to go back into breeding harems. This procedure normally works very well. The juveniles seem to enjoy having the adults in their presence, and the adults are certainly under less stress than they would be going directly into breeding harems with other adult animals. Once they have recovered, it is much easier to introduce them back into groups. Normally, individual females or small groups of females are placed together to make up totally new harems.
The second procedure to be described is our foster infant program. Approximately 30 infants a year are either rejected or orphaned at or near birth. We have no nursery, and we have a policy which prohibits taking those infants home and hand rearing them. An alternative is fostering as a means of saving these offspring. Our technique is to take the individual infant out to the colony and go from crib to crib to locate females that have a particular interest in the infant. We then select one of the females carrying an infant of a similar age, place them in an individual cage, and introduce the infant to be fostered to her. Normally the female will accept the infant. If everything seems to be going well, we leave all 3 animals inside for a 5 to 7 day period for observation before returning them to the harem. Occasionally it's necessary to repeat this procedure with another female if incompatibility develops. In some cases, if lactation doesn't appear to be adequate at first, we supplement the infant with infant formula 2 to 3 times a day until we are sure that it is feeding adequately.
The third procedure is introduction of breeder males into groups of established breeder females. At the time of quarterly TB testing, animals are palpated for pregnancy. Once we finish testing the colony, we evaluate conception rates in the various harems, and develop a plan to share the more productive males with those harems that have lower conception rates. We accomplish this sharing by swapping the groups of females, leaving the males in place. We find that we have much better acceptance of the males by using this technique. We also have found that it is more successful when we select groups of females from distant parts of the colony where they have not had visual and vocal contact with one another. Normally we can determine the compatibility very quickly. When it is not successful, we generally remove the most aggressive females from the group. Other times it is necessary to split up the group completely and establish new breeding harems.--Steve Pearson, Hazleton--Texas Primate Center, P.O. Box 549, Alice, TX 78332.
California Primate Research Center
The problem I want to address today is inbreeding, a problem for the management of all captive primate populations where natural male migration is not possible. Our population consists of approximately 1000 rhesus, housed in 1/2 acre field cages at the California Primate Research Center (CPRC). David Glenn Smith, using Paternity Exclusion Analysis at the CPRC, found that rhesus mate randomly with regard to paternal kinship, but only one case (full sibs) of matrilineal inbreeding was found. In an earlier study (1977-84), Smith compared the 40 inbred individuals he found in the CPRC field cages to non-inbred individuals. The only statistically significant difference was that inbred animals had slightly lower birth (but not growth) rates. However, he noted that reproductive loss is greater after 3-4 generations, so there may be a problem in the future at the CPRC. Inbreeding is a problem in biomedical research when a source population becomes genetically homogeneous and distinct from other primate center populations. (Are research results compatible if test animals come from genetically distinct populations?)
The CPRC has been working on two solutions to the problem of inbreeding. The first is to increase genetic diversity through the introduction of rhesus from diverse geographical populations. Most of the CPRC rhesus are of Indian origin. In 1982, 3 Chinese rhesus males were introduced into a cage of 53 juvenile Indian rhesus. These males have now produced a number of hybrids who weren't significantly different from non-hybrids when they were studied as infants by D. G. Smith et al, but which now appear larger than their Indian counterparts.
In October, 1987, we introduced 3 Burmese rhesus males into a field cage of 73 juveniles (1.5 to 3.5 years old). At that time, we were concerned about trauma due to general xenophobia, and the fact that some of the Chinese hybrid juvenile males were actually larger than the Burmese males. However, when the Burmese males were released into the field cage, all juveniles retreated to the ceiling and sides of the cage, where they stayed for several days. We had one breeding season (1987-88) with the Burmese males in the field cage, resulting in 13 conceptions and 11 live births to date. All the mothers are primiparous and are doing beautifully, but the Burmese males did not fare so well. In January 1988, the oldest of the males (approximately 16 years) was hospitalized for dehydration and hypothermia. In March, the alpha male (second oldest at approximately 12 years) was badly traumatized by unknown assailants and removed from the cage. Neither male was returned to the field cage. By late spring, the new alpha male was a 3-year-old Chinese hybrid, even though there were 6 older males in the cage with him. Upon completion of Dave Smith's paternity exclusion analysis we should know if the fathers of the 1988 infants in this field cage are Burmese and/or the younger Chinese and Indian males.
The second solution is an infant cross fostering program launched by Steve Smith in 1984, and first reported at the ASP meetings in Austin in 1986. This program started as a series of infant swaps between field cages, and Steve Smith was able to make 36 swaps in 3 years with a 75% success rate. Thirty-two of the 36 fostered animals are still in the field cages, including one 5-year-old female (who unfortunately had a stillbirth this year), and eight 4-year-old males. We don't know yet what their reproductive success (RS) will be.
This year to date we have been successful in 11 of 14 infant foster attempts (79%). The three failures were an infant reject, a mutual reject, and a maternal reject. Examining both infant and maternal behavior, it looks as though we need to pay more attention to temperament differences in order to increase fostering success rates. Infants who are highly agitated during the transfer process may not be very good candidates for fostering; the best foster pairs are very calm mothers with very calm infants. Six of the 11 fosters this year were genetic fosters--that is, we took either full Chinese, 1/2 Chinese, or 1/2 Burmese rhesus infants born indoors and swapped them with infants in the field cages. These fosters were all successful. We did just one straight foster of an animal that was born in the field cages. He was rejected by his mother (who had been quite ill), raised in the nursery for nearly a month, and then fostered to a primiparous female who had a stillbirth. Both mother and infant are doing well.
It has also become apparent from D. G. Smith and S. Smith's work at the CPRC that both males and females inherit maternal rank but 1) RS of natal males is significantly correlated with maternal rank and not paternal rank (although the son of a high-ranking female and an alpha male has an additional RS bonus) and 2) the advantage of maternal rank does not seem to accrue to daughters. Given greater variance in male RS, we are now attempting to place male foster infants only on high-ranking (especially alpha) females, and placing female foster infants only when male infants are not available.
There are several things we don't know about inbreeding but are working to understand. One is the actual lifetime RS of fostered individuals; we're banking on very successful male breeders in the future. We don't know enough about the long term effects of inbreeding as we haven't had enough inbred animals to know how serious a problem this will be. We also don't have enough information about hybrid vigor, although our subjective impression is that Chinese hybrid males seem consistently larger and higher ranking than their pure Indian counterparts. In the future we will continue the infant cross fostering program and attempt to diversify the CPRC rhesus population even further. We are expecting some Vietnamese animals from Sukhumi this fall and hope to use these animals to start a new field cage with juvenile rhesus of Indian stock--Linda M. Scott, California Primate Research Center, Davis, CA 95616.
-------------------------------------------------------------------
This is a transcript of a workshop presented at the 11th meeting of the American Society of Primatologists, June 2-5, 1988, at Loyola University, New Orleans, LA. The workshop was organized, and the transcript edited, by Margaret R. Clarke, Delta Regional Primate Research Center of Tulane University, Three Rivers Road, Covington, LA 70433.
* * *
IPS
The XIIIth Congress of the International Primatological Society will be held in Japan, from July 18-24, 1990. Registration, papers, poster sessions, and audiovisual sessions will take place in Nagoya July 18-21, while symposia will be held July 23-24 in Kyoto. Persons interested in organizing symposia should send their proposals to Professor Toshisada Nishida, Dept. of Zoology, Faculty of Science, Kyoto Univ., Kitashirakawa-Oiwakecho, Sakyo-ku, Kyoto 606, Japan, by May 1, 1989. Address all other correspondence concerning the meeting to Professor Osamu Takenaka, Primate Research Institute, Kyoto Univ., Kanrin, Inuyama, Aichi 484, Japan.
Fertility in Great Apes
A three day meeting will be held in Atlanta, GA on June 15-17, 1989, organized by Yerkes Regional Primate Research Center of Emory University, and cosponsored by Zoo Atlanta and the National Zoo. Contact Dr. Kenneth G. Gould, Yerkes Regional Primate Research Center, Emory University, Atlanta, GA 30322 [404-727-7720].
NABR
The National Association for Biomedical Research will hold a conference on federal legislation April 30-May 2, 1989, at the Capitol Hilton in Washington, DC, to "inform our members and educate the 101st Congress." Representatives from both the House and Senate will be invited to explain how the animal research issue is viewed from their perspectives. Panel discussions, strategy planning, and the opportunity to establish or improve relationships with Congressional offices through individual visits to Capitol Hill are also on the agenda. The address of NABR is 818 Connecticut Avenue N.W., Suite 303, Washington, DC 20006 [202-857-0540].
PHS Policy Workshop
Another workshop on the implementation of Public Health Service policy on laboratory animals, sponsored by the Office for Protection from Research Risks of NIH, will be held on February 9-10, in Salt Lake City, UT. These workshops are open to institutional administrators, members of animal care and use committees, laboratory animal veterinarians, investigators, and those with responsibility for institutional animal care and use programs. Contact Joan Provost, Conferences and Institutes, Univ. of Utah, Salt Lake City, UT 84112 [801-581-5809].
ASP
The twelfth meeting of the American Society of Primatologists, hosted by the University of South Alabama, will be held August 27-30, 1989. Paper sessions, symposia, posters, exhibits, and business meetings will take place at the Riverview Plaza Hotel in Mobile, AL. Committee meetings, including the Executive Committee meeting, are scheduled for Sunday afternoon, August 27. An evening reception for all registrants will follow. Scientific paper presentations will begin Monday morning, August 28, with the final session ending about noon on Wednesday, August 30. For additional information contact Mike Raleigh, Program Chair, at the Department of Psychiatry, University of California, Los Angeles, CA 90024 [213-825-0565].
* * *
No Controversy
The opening paragraph in the article "Chimps and Research: Endangered?" (Laboratory Primate Newsletter, 1988, 27[4], 9-12) states that scientists in the conservation and biomedical communities are "at each others' throats," the latter "clamoring for chimps for AIDS research." On the next page there is a reference to "a letter to Nature " by Alfred Prince and Jane Goodall, opposing importation of chimpanzees from the wild for research and arguing that the current captive population is sufficient for research needs.
The letter mentioned is a position paper by Alfred M. Prince, Jan Moor-Jankowski, Jorg W. Eichberg, Huub Schellekens, Rudolph F. Mauler, Marc Girard, and Jane Goodall, which appeared in volume 333[6173], 1988, p. 513 of Nature. Not only Dr. Prince but five of the other co-authors are well-known for their biomedical research using chimpanzees, so our paper proves that there is no controversy between us and our co-author, Dr. Jane Goodall, the noted conservationist. Dr. Eichberg, the co-discoverer of the chimpanzee as a model for human AIDS, is responsible for a large chimpanzee colony in the U.S., as am I. Dr. Schellekens directs the hepatitis and AIDS program in the large Dutch chimpanzee center, supported by the European Economic Community as an international resource, while Dr. Girard directs the vaccine research at Pasteur Vaccins, France, and Dr. Mauler the vaccine program at Behringwerke, West Germany.--J. Moor-Jankowski, Director, Laboratory for Experimental Medicine and Surgery in Primates, New York University School of Medicine, 550 First Avenue, New York, NY 10016.
Restrictions on Use
Since the "endangered" classification places significantly greater restrictions on the use of such animals, including those already in captivity, many areas of disease prevention and treatment research, including studies on AIDS, could be severely hampered by this reclassification, particularly those for which chimpanzees may be the only appropriate research animals. Any reclassification should be based upon a responsible and objective survey of the number and circumstances of chimpanzees in the wild and not on anyone's guess or wish. It is important to note that no chimpanzees are being imported to the United States from their natural habitats in the wilds of Africa.
In 1973, the U.S. joined the Convention on International Trade in Endangered Species (CITES), and it became law in 1975. Since then, the U.S. has not imported chimpanzees from the wild. Thus, reclassifying the chimpanzees as "endangered" would not provide additional protection of these animals from exportation from Africa to the U.S., since our government already abides by CITES. And, contrary to current rumors, the National Institutes of Health does not plan importation of wild chimpanzees.
It should also be recognized that the NIH has established a National Chimpanzee Breeding Program in the U.S. to insure that chimpanzees do not become extinct in captivity and that there will remain an adequate supply for biomedical research in this country. This Program and the concern of the scientific and zoological community can assure the long-term survival of this species in captivity. Regardless of how chimpanzees are classified, it is essential for the advancement of several areas of medical studies that those chimpanzees presently in research institutions and their progeny of the future continue to be available for research so long as their numbers in research and breeding establishments in the U.S. are stable or increasing.--Frederick A. King, Director, Yerkes Regional Primate Research Center, Emory University, Atlanta, GA 30322.
Pet Chimpanzees
We know the number of chimpanzees in research institutions and government facilities, and even have a general idea of the number left in the wild, but how can we account for the number in this country owned by private individuals as pets? The Primate Supply Information Clearinghouse (PSIC) has received calls from owners of fairly young (2-3 years old) solitary chimps who want to locate and buy another. Many of these pet owners are upset at the problems locating a supply, and if they do find some breeder who has one for sale, they don't like the current price! Would changing the status from Threatened to Endangered essentially eliminate the private ownership of young chimpanzees? How many of these privately held animals will have to be "given" to a colony or sanctuary when they become adults? Who is monitoring the health status and census of these animals, and how did they become "obtainable" by anyone with the money? I doubt if there are answers to these questions, and I doubt if we can investigate very thoroughly, as I have had numerous dealings with commercial importers, distributors, and pet stores selling these and other "exotic" animals. Their records, histories, and dates of receipt become conveniently vague. There are also problems with which authorities to report these animals to, lack of manpower among the authorities, and lack of somewhere to send confiscated animals.--Cathy A. Johnson-Delaney, PSIC, University of Washington, Seattle, WA 98195.
Compromise
The U.S. Fish and Wildlife Service has decided to reclassify wild chimpanzees as "endangered," while leaving chimps living in research facilities under the less dire heading of "threatened." The compromise will allow experimentation on man's closest relative to continue, while bringing into play more stringent restrictions on the importation of chimpanzees from Africa, where the species is under severe pressure due to massive habitat destruction, hunting, and capture.--From a News and Comment article in Science, December 2.
* * *
AIDSLINE
The National Library of Medicine has added a new database to its MEDLARS family--AIDSLINE. NLM and the NIH Office of AIDS Research recently announced the availability of this database which contains some 13,000 references to scientific articles about AIDS. Clinical and research aspects of the disease, epidemiology, and health policy issues are covered. AIDSLINE will be updated with the addition of 200-300 records twice each month.
To make individual online searching easier, NLM offers GRATEFUL MED, a microcomputer-based software that provides a user-friendly approach to searching. Telephone inquiries about AIDSLINE, GRATEFUL MED, and user codes may be made to NLM's Management Section, 800-638-8480.--From the Wisconsin Regional Primate Research Center's Primate Library Report, 1988, 16.
Animal Welfare Information Center
The Animal Welfare Information Center at the National Agricultural Library has recently completed two new publications: "Training Materials in the Collections of the National Agricultural Library Relating to Animal Welfare" and "Audio-Visuals in the Collections of the National Agricultural Library Relating to Animal Welfare." The publications are available to the general public through local libraries. For further information, contact Lending Branch, NAL, Beltsville, MD 20705 [301-334-3755].
PSIC
"New Listings," the weekly bulletin put out by the Primate Supply Information Clearinghouse, which serves as a communication link between researchers, breeding colonies, laboratories, and information services involving nonhuman primates, is undergoing some changes. Ads are now run weekly for 4 weeks, with optional extensions. In addition, there is a new bulletin, "Continuing Listings," containing long-term programs, which accompanies the first regular issue of "New Listings" each month.
Dr. Cathy A. Johnson-Delaney, the PSIC Coordinator, invites long-term programs, including breeding colonies, which may provide or need animals, tissues, services, or equipment, to list in "Continuing Listings," as well as listing their periodic needs or availabilities in "New Listings." Contact Dr. Johnson-Delaney at the Primate Information Center, Regional Primate Research Center, SJ-50, University of Washington, Seattle, WA 98195 [206-543-5178].
* * *
Simian immunodeficiency virus (SIV) belongs to the family Retroviridae (subfamily Lentivirinae) and is closely related to human immunodeficiency virus types 1 and 2 (HIV-1 and HIV-2), the etiologic agents of acquired immunodeficiency syndrome (AIDS). Although no reports of infection in humans have been documented, the expanding use of SIV as a model of HIV infection has raised concern about the potential risk of SIV transmission to humans. Therefore, a working group was established by CDC and the National Institutes of Health (NIH) to formulate specific guidelines intended to minimize the risk of SIV transmission to humans.
Background
Originally reported in 1985, the first isolate from a rhesus macaque was called simian T-lymphotropic virus III (STLV-III) (Daniel et al.). Viral isolates have since been obtained from several species of nonhuman primates including African green monkeys (Ohta et al., 1988), sooty mangabeys (Fultz et al., 1986), pig-tailed macaques (Benveniste et al., 1986), and stump-tailed macaques (Lowenstine et al., 1987). Limited studies of wild-caught African green monkeys from Central Africa indicate a seroprevalence of approximately 30-50%, apparently without associated immunodeficiency disease. The seroprevalence of SIV among rhesus monkeys in captive colonies (whether from natural infections or interspecies transmission) appears to be low (i.e., 0-1%) (Daniel et al., 1988). In contrast, captive sooty mangabeys may have seroprevalence rates as high as 80% (McClure, personal communication). The prevalence of SIV infection among many other nonhuman primate species is unknown.
In rhesus monkeys and other susceptible nonhuman primate species (e.g., pig-tailed macaque, crab-eating macaque), SIV infection leads to a chronic wasting disease syndrome with depletion of CD4 (T4) lymphocytes and lymphadenopathy. The clinical course of this infection in monkeys, like that of AIDS in humans, is complicated by opportunistic infections (King, 1986). SIV also causes a primary encephalopathy with many of the features of HIV-associated encephal- opathy (Ringler et al., 1988). Therefore, SIV infection is an important animal model of AIDS.
SIV proteins, especially the viral core proteins (i.e., p24, capsid protein), are antigenically related to HIV-1 proteins (Kanki et al., 1985). Some SIV isolates, however, are antigenically more related to HIV-2 than to HIV-1 by cross-reactivity of viral capsid and envelope proteins. SIV isolates that have been molecularly cloned share approximately 75% of their genomic sequences with HIV-2 and approximately 30% with HIV-1 (Chakrabarti et al., 1987). SIV isolates are clearly distinct from Type D primate retrovirus (i.e., simian retrovirus 1) that also causes a form of chronic wasting immunodeficiency disease in several primate species (Heidecker et al., 1987). Also, SIV is distinct from simian T-cell lymphotropic virus type I (STLV-I), which shares extensive genomic sequences with human T-lymphotropic virus type I and is associated with T-cell lymphomas in nonhuman primates (Watanabe et al., 1985).
SIV can be isolated from a variety of tissues and body fluids``including blood, plasma, cerebrospinal fluid, and parenchymal tissues``of infected nonhuman primates. Limited data exist concerning the presence or concentration of virus in semen, cervical secretions, saliva, urine, breast milk, and amniotic fluids of experimentally or naturally infected nonhuman primates. However, the virus apparently is rarely isolated from semen, urine, and saliva despite repeated attempts at isolation (M. Daniel, N. Lerche, personal communication). There is no evidence to indicate that SIV is transmitted by the respiratory route (N. Lerche, H. McClure, M. Daniel, personal communication).
The cell tropism of SIV in culture depends partially on the strain of virus propagated and conditions of cell culture. Strains of SIV have been successfully cultured in human lymphocyte cell lines (e.g., HuT 78, HT, CEMx174) and in primary human and nonhuman primate peripheral blood leukocyte cultures (Kannagi, Yetz, & Letvin, 1985). SIV appears to be primarily tropic for CD4 (T4)-positive leukocytes and has not been successfully propagated in B-lymphocyte cell lines (e.g., Raji). SIV antigen has been demonstrated by immunohistochemical methods in lymph node sinus histiocytes, macrophages, and giant cells (Ward et al., 1987), as well as in macrophage-derived cells in brain tissue from diseased monkeys (Ringler et al., 1988).
Limited data exist concerning the reactivation of Herpesvirus simiae (B virus) or other latent infectious agents in SIV-infected macaque monkeys. However, all macaque monkeys not proven to be free of B virus infection, regardless of SIV infection status, should be regarded as infected with B virus and handled according to published guidelines (CDC, 1987). The routine screening of macaques for evidence of B virus infection or SIV infection is not recommended. However, in situations in which studies may cause immunosuppression (e.g., during experimental SIV infections), the investigator may elect to determine the infection status of the animals because B virus shedding may be enhanced in infected animals.
Evaluating the Risk of SIV Transmission to Humans
The risk, if any, of human infection with SIV has not been defined. However, since SIV shares many characteristics with HIV, many of the same biosafety precautions are indicated. No serologic or virologic evidence of infection in humans exists; specific precautions in handling SIV are based on recommendations developed for HIV and other lentiviruses. No licensed tests exist for serologic evaluation of humans exposed to SIV, which complicates medical surveillance and investigations of the virus infection following exposure to SIV. In addition, the antigenic cross-reactivity between SIV and HIV may complicate testing of exposed humans.
However, standardized serologic procedures that test for SIV antibody are used in laboratories performing research with the virus. Recently, a protein unique to SIV and HIV-2 (product of gene vpx) was used as antigen in a serologic test that may allow easier distinction between HIV-1 and SIV antibodies (Henderson et al., 1988). Furthermore, gene amplification (i.e., polymerase chain reaction) may allow differentiation of specific virus gene sequences directly from specimens obtained from exposed persons. Based on these events, development of specific and sensitive tests is under way.
Guidelines to Prevent SIV Infection in Laboratory Workers and Animal Handlers
Exposure Concerns. In the laboratory, SIV must be presumed to be present in all SIV cultures, in all materials derived from such cultures, in all specimens from SIV-antibody-positive nonhuman primates, and in/on all equipment and devices coming in contact with these materials. In this setting, the skin (especially when scratches, cuts, abrasions, dermatitis, or other lesions are present) and mucous membranes of the eye, nose, and mouth should be considered as potential pathways for virus entry; contact of these sites with SIV-containing materials should be considered an exposure to SIV.
Biosafety Levels. Biosafety level (BSL) 2 standards and special practices, containment equipment, and facilities, as described in the CDC/NIH publication Biosafety in Microbiological and Biomedical Laboratories (1984), are recommended for activities involving all clinical specimens, body fluids, and tissues from SIV-infected primates. In laboratories maintaining BSL 2, personnel must have documented specific training in handling primate retroviruses, and the laboratory must have limited and properly secured access and written standard operating procedures for techniques in which SIV is used. Procedures involving cultures of SIV should be conducted in biological safety cabinets or other physical containment equipment.
Inoculation Precautions. In the research laboratory, inoculation of SIV-containing material represents an important potential route of exposure to SIV in humans. The use of syringes, needles, glass, and other sharp objects should be avoided, but when their use is essential, needles and disposable cutting instruments should be discarded after use into a lidded puncture-resistant container located in the work area. Needles should not be resheathed, bent, broken, removed, or otherwise manipulated by hand.
Gloves. Latex or vinyl gloves should be worn by all personnel engaged in activities that may involve direct skin contact with infectious specimens, cultures, or tissues. Gloves should not be washed or disinfected for reuse; reuse of such gloves may cause "wicking" (i.e., enhanced penetration of liquids through undetected holes in the glove) or deterioration of the gloves. When gloves have become visibly contaminated, they should be carefully removed and, after the hands are washed, replaced with a fresh pair of gloves. Handwashing with soap and water immediately after infectious materials are handled and work is completed, even when gloves have been worn, should be routine practice.
Clothing. Laboratory coats, gowns, or uniforms should be worn by laboratory workers when engaged in any work involving SIV or materials known or suspected to contain SIV. Clothing that becomes contaminated with SIV or SIV-containing materials should be decontaminated before being laundered or discarded. Clothing can be decontaminated by extensive soaking of the garment with chemical disinfectants (e.g. 1 to 10 dilution of household bleach).
Aerosol Control. Although aerosol transmission of SIV has not been demonstrated, the generation of aerosols, droplets, splashes, and spills should be avoided. A biological safety cabinet should be used for all procedures that might generate aerosols or droplets and for all infected cell culture manipulations. When centrifuging infected materials, centrifuge containers with safety caps that are designed to contain aerosols (in the event of spillage) should be used. When cell sorters are used, plastic shielding or other containment devices should be used to reduce exposure to infectious droplets.
Contaminated Virus Preparations. During the propagation of SIV in the research laboratory, manipulation of concentrated virus preparations or conduct of procedures that may produce aerosols or droplets should be performed in a BSL 2 facility, with additional practices and containment equipment recommended for BSL 3 (CDC, 1984). These practices should include wearing closed-front surgical-type gowns, masks and protective eyewear or face shields, and latex or vinyl gloves that extend to cover the wrist and sleeves of the surgical gown. Activities involving large-volume production or manipulation of highly concentrated SIV should be conducted in a BSL 3 facility, using only BSL 3 practices and equipment. All discarded cultures and laboratory supplies used in experimental manipulations of cultures should be autoclaved before disposal.
Decontamination. The susceptibility of SIV to chemical disinfectants has not been defined. Work surfaces, however, should be decontaminated daily with commercially available chemical disinfectants such as sodium hypochlorite solution 10% (1 to 10 dilution of household bleach), ethanol 70-85%, or ethanol-iodine complex 2%. These effectively inactivate HIV (Martin, McDougal, & Loskoski, 1985; Resnick et al., 1986). Prompt decontamination of spills (immediate absorption and control of the spill and soaking of the contaminated area with chemical disinfectant) should be standard practice. Gloves should be worn when cleaning up such spills. The use of plastic-backed absorbent padding to control spillage during manipulation of cultures or other SIV-containing fluids should be encouraged.
Animal Biosafety Levels. Animal BSL 2 practices, containment equipment, and facilities are recommended for activities involving nonhuman primates or any animals experimentally infected or inoculated with SIV. Animal-care personnel, investigators, technical staff, and other persons who enter animal rooms should wear coats, protective gloves, coveralls or uniforms, and, as appropriate, face shields or surgical masks and eye shields to protect the skin and mucous membranes of the eyes, nose, and mouth.
Handling SIV-Infected Nonhuman Primates. Nonhuman primates experimentally infected with SIV may have other primary, as well as opportunistic, pathogens in their body fluids and tissues. Thus, laboratory workers and animal handlers should follow accepted BSL 2 practices at all times to prevent inadvertent exposure to agents that may be present in clinical specimens or body fluids. All macaque monkeys not known to be free of Herpesvirus simiae (B virus) should be regarded as infected and handled according to published guidelines (CDC, 1987).
Personnel Training. Primary investigators, other scientific personnel, and other persons who handle nonhuman primates used in SIV research should be trained in proper methods of animal restraint and use of protective clothing. Animal handlers should be familiar with various drugs routinely used for providing chemical restraint and with proper procedures for administering medications. All persons engaged in research involving nonhuman primates should be specifically trained in the natural history of SIV infection. Particular attention should be given to the need to wear protective clothing to prevent mucous membrane contact with potentially infectious material, particularly animal blood from an SIV-infected nonhuman primate. Caution should be emphasized during venipuncture procedures or the administration of injections to nonhuman primates involved in SIV research. Intravenous injections of nonhuman primates should be done while the animal is anesthetized and should be administered through a plastic or teflon catheter with syringes fitted with interlocking connectors.
Medical Surveillance. A licensed test specific for SIV antibody is not yet available. Standardized serologic procedures to identify SIV antibody are used in laboratories performing research with the virus. A medical surveillance program should be in place in all laboratories that test specimens, conduct research, or produce reagents involving SIV. The nature and scope of the surveillance program will vary by institutional policy and applicable local, state, and federal regulations (U.S. Departments of Labor & of Health and Human Services, 1987). Laboratories performing research with SIV should initiate a program to store serum from laboratory workers. Serum specimens should be collected at 6-month intervals and stored. Routine testing of the serum is optional but, if performed, should be done using standardized serologic procedures in qualified laboratories.
Human Exposure to SIV. If a laboratory worker has a parenteral, skin, or mucous membrane exposure to blood, body fluids, or virus culture material from nonhuman primates, the source material should be identified and, if possible, tested for the presence of SIV. All wounds incurred from SIV-infected nonhuman primates or from SIV-contaminated instruments hould be cleansed with soap and water. Such incidents should be reported to the animal-care supervisor and/or laboratory supervisor and recorded in an accident report log. If the source material is positive for SIV antibody, virus, or antigen, or unavailable for examination, the worker should be counseled regarding the risk of infection and evaluated medically. The worker should be advised to report and to seek medical evaluation for any acute febrile illness that occurs within 12 weeks after the exposure. Medical evaluation should include examination for serum antibody against SIV. Seronegative workers should be retested 6 weeks after the exposure and periodically thereafter (e.g., 12 weeks and 6 months after exposure). All institutions should establish written policies regarding the management of laboratory exposure to SIV; such policies should deal with confidentiality, counseling, and other related issues. The lack of data concerning the potential transmission of SIV between humans does not allow for specific recommendations concerning behavior changes in a person confirmed seropositive for SIV. However, an SIV-positive person should not donate blood.
Postexposure Treatment. No effective prophylactic treatment for SIV exists; research is needed in animals concerning postexposure prophylaxis (e.g., immune globulin, antiviral therapy). Data from such research may be useful in future exposures of humans to SIV. -- [From Morbidity and Mortality Weekly Report, 37, 1988, 693-694, 699-704.]
References ;
Benveniste, R. E., Arthur, L. O., Tsai, C.-C., Sowder, R., Copeland, T. D., Henderson, L. E., & Oroszlan, S. (1986). Isolation of a lentivirus from a macaque with lymphoma: comparison with HTLV-III/LAV and other lentiviruses. Journal of Virology, 60, 483-490.
CDC (1984). Biosafety in microbiological and biomedical laboratories. Bethesda, MD: US Dept of Health & Human Services, Public Health Service, DHHS publication no. (CDC)84-8395.
CDC (1987). Guidelines for prevention of Herpesvirus simiae (B virus) infection in monkey handlers. Morbidity and Mortality Weekly Report, 1987, 680-682, 687-689. [Reprinted in Laboratory Primate Newsletter, 1988, 27[1], 9-12.
Chakrabarti, L., Guyader, M., Alizon, M., Daniel, M. D., Desrosiers, R. C., Tiollais, P., & Sonigo, P. (1987). Sequence of simian ac- quired immunodeficiency virus from macaque and its relationship to other human and simian retroviruses. Nature, 328, 543-547.
Daniel, M. D., Letvin, N. L., King, N. W., Kannagi, M., Sehgal, P. K., Hunt, R. D., Kanki, P. J., Essex, M., & Desrosiers, R. C. (1985). Isolation of T-cell tropic HTLV-III-like retrovirus from macaques. Science, 228, 1201-1204.
Daniel, M. D., Letvin, N. L., Sehgal, P. K., Schmidt, D. K., Silva, D. P., Solomon, K. R., Hodi, F. S. Jr., Ringler, D. J., Hunt, R. D., King, N. W., & Desrosiers, R. C. (1988). Prevalence of antibodies to three retroviruses in a captive colony of macaque monkeys. International Journal of Cancer, 41, 601-608.
Fultz, P. N., McClure, H. M., Anderson, D. C., Swenson, R. B., Anand, R., & Srinivasan, A. (1986). Isolation of a T-lymphotropic retrovirus from naturally infected sooty mangabey monkeys (Cercocebus atys). Proceedings of the National Academy of Science, 83, 5286-5290.
Heidecker, G., Lerche, N. W., Lowenstine, L. J., Lackner, A. A. Osborn, K. G., Gardner, M. B., & Marx, P. A. (1987). Induction of simian acquired immune deficiency syndrome (SAIDS) with a molecular clone of a type D SAIDS retrovirus. Journal of Virology, 61, 3066-3071.
Henderson, L. E., Sowder, R. C., Copeland, T. D., Benveniste, R. E., & Oroszlan, S. (1988). Isolation and characterization of a novel protein (X-ORF product) from SIV and HIV-2. Science, 241, 199-201.
Kanki, P. J., McLane, M. F., King, N. W. Jr., Letvin, N. L., Hunt, R. D., Sehgal, P., Daniel, M. D., Desrosiers, R. C., & Essex, M. (1985). Serologic identification and characterization of a macaque T-lymphotropic retrovirus closely related to HTLV-III. Science, 228, 1199-1201.
Kannagi, M., Yetz, J. M., & Letvin, N. L. (1985). In vitro growth characteristics of simian T-lymphotropic virus type III. Proceedings of the National Academy of Science, 82, 7053-7057.
King, N. W. (1986). Simian models of acquired immunodeficiency syndrome (AIDS): A review. Veterinary Pathology, 23, 345-53.
Lowenstine, L. J., Lerche, N. W., Marx, P., Pederson, N. C., Jennings, M., & Gardner, M. B. (1987). Update on simian immunosuppressive retrovirus (SIV) studies at the California Primate Research Center (CPRC). Presented at the Symposium on Nonhuman Primate Models for the Acquired Immunodeficiency Syndrome (AIDS), Atlanta, GA, October 8-10.
Martin, L. S., McDougal, J. S., & Loskoski, S. L. (1986). Disinfection and inactivation of the human T lymphotropic virus type III/lymphadenopathy-associated virus. Journal of Infectious Disease, 152, 400-403.
Ohta, Y., Masuda, T., Tsujimoto, H., Ishikawa, K., Kodama, T., Morikawa, S., Nakai, M., Honjo, S., & Hayami, M. (1988). Isolation of simian immunodeficiency virus from African green monkeys and seroepidemiologic survey of the virus in various nonhuman primates. International Journal of Cancer, 41, 115-122.
Resnick, L., Veren, K., Salahuddin, S. Z., Tondreau, S., & Markham, P. D. (1986). Stability and inactivation of HTLV-III/LAV under clinical and laboratory environments. Journal of the American Medical Society, 255, 1887-1891.
Ringler, D. J., Hunt, R. D., Desrosiers, R. C., Daniel, M. D., Chalifoux, L. V., & King, N. W. (1988). Simian immunodeficiency virus-induced meningoencephalitis: Natural history and retrospective study. Annals of Neurology, 23(suppl), S101-S107.
Ward, J. M., O'Leary, T. J., Baskin, G. B., Benveniste, R., Harris, C. A., Nara, P. L., & Rhodes, R. H. (1987). Immunohistochemical localization of human and simian immunodeficiency viral antigens in fixed tissue sections. American Journal of Pathology, 127, 199-205.
Watanabe, T., Seiki, M., Tsujimoto, H., Miyoshi, I., Hayami, M., & Yoshida, M. (1985). Sequence homology of the simian retrovirus genome with human T-cell leukemia virus type I. Virology, 144, 59-65.
U.S. Department of Labor, U. S. Department of Health and Human Services. (1987). Joint advisory notice: HBV/HIV. Federal Register, 52 [210], 41818-41824.
* * *
Pulmonary Hypertension
A newly discovered animal model for pulmonary hypertension, based on naturally occurring pulmonary vascular disease in the stumptailed macaque (Macaca arctoides) , may be helpful in developing biochemical markers for early detection of the disease in humans, according to Dr. Kenneth M. Weesner of the Bowman Gray School of Medicine in Winston-Salem, NC. An appropriate animal model for human pulmonary vascular disease is critically needed, because diagnosis often comes after the disorder is far advanced and perhaps irreversible. The cause of primary pulmonary hypertension is completely unknown. This disease is progressive and eventually causes severe symptoms, although it may be clinically silent for many years.
A study of the pulmonary circulation of stumptailed macaques revealed that nearly one-third of 63 animals had severe medial thickening of pulmonary arteries that was not due to a congenital heart defect. Hemodynamic studies of 29 live animals of different ages revealed that 8 had elevated pulmonary artery pressure, 10 had normal pressure, and 11 had intermediate pulmonary pressure. Those with elevated pulmonary artery pressure had a blunted increase in pulmonary pressure compared to those with normal pressure, when exposed to hypoxic conditions which cause constriction of the pulmonary artery. The pulmonary hypertension seen is not severe enough to warrant trials of therapeutic measures.
"The most crucial research question involving this colony is a biochemical marker. There is some evidence that platelet activation may lead to the increased pulmonary artery pressure that we are seeing, both in monkeys and in humans," Dr. Weesner says. "This may serve as a biological marker that we can use in the early stages of the disease to identify people who can't be identified hemodynamically or by echocardiography."
If vasoconstriction is involved, it may be possible to develop biological and biochemical markers for pulmonary hypertension using various vasoactive mediators. Echocardiography can be used to detect pulmonary hypertension in humans, but because the disease may be silent the dilemma of which asymptomatic patients to test remains. Biological markers also may make the stumptailed macaque model useful for evaluating prospective treatments for pulmonary hypertension.--From a report by Jonathan Adams in the Research Resources Reporter, November, 1988, 12[11], 9-10.
False-Positive AIDS Tests
The enzyme-linked immunosorbent assay, ELISA, a serum screening test widely used for detection of antibodies to HIV, the human immunodeficiency virus that causes acquired immune deficiency syndrome (AIDS), can produce a high rate of false-positive results in testing certain species of African monkeys. Researchers at the California Regional Primate Research Center (PRC) in Davis suggest that a reaction between natural antibodies and cellular protein contaminants of preparations of viral antigens might be responsible for the high rate of false-positive results and may mask some true-positives.
"ELISA testing is a good method of screening for antibodies, but it is imperative to verify positive ELISA results with a test that is more specific, such as the Western blot," says Dr. Niels C. Pedersen.
Approximately one-fourth of the sera from 526 old-world primates representing 50 different species of monkeys tested positive by the ELISA for antibodies to 3 different retroviruses. The viruses were human T-lymphotropic virus I (HTLV-I), which causes T cell leukemia; HIV; and simian retrovirus type I (SRV-1, a type D retrovirus that also causes simian AIDS). This high incidence of positive responses prompted the investigators to retest the ELISA positives by the Western blot method. Only 20% of those retests were positive.
Six species had the highest concentration of false-positive reactions: Mandrillus sphinx (mandrill), Macaca fascicularis (crab-eating Macaque), M. silensus (lion-tailed macaque), Cercopithecus aethiops (African green monkey), Cercopithecus neglectus (DeBrazza's guenon), C. cephus (moustached guenon), and Miopithecus talapoin (talapoin). Patterns of false-positive reactions were identical in the six species. In other tests, the sera reacted positively to nonretroviruses and cellular debris, suggesting that the sera reacted with cellular material that contaminated the viral protein preparations.
"Some species of animals have a lot of nonspecific antibodies, and it is only by doing the next step, Western blot testing, that you can determine if the ELISA positives are specific or nonspecific," says Pederson. He adds that Western blot testing, although more accurate than ELISA and other available tests, is partly subjective and has quality-control limitations. He feels that more specific and objective approaches need to be developed for even better retrovirus antibody testing.--From a report by Barbara J. Proujan in the Research Resources Reporter, September, 1988, 12[9], 9-12.
* * *
Victor E. Collazo
Caribbean Primate Research Center
The Caribbean Primate Research Center maintains a large number of rhesus monkeys (Macaca mulatta) in harem social groups. Several times, the sires have shown paternal behavior towards their own offspring in these social groups, going as far as temporarily adopting infants. The sire E411, a 12-year-old male, housed with 3 adult females and 2 yearlings in a 40 sq-m concrete pen, expressed this paternal behavior beyond the limits of its own progeny.
One morning, a caretaker noticed that E411 was carrying a live infant. To his surprise, after closer observation, the employee discovered that the "infant" was not a monkey, but an approximately three-week-old cat which apparently had wandered inside the corral and was caught (or adopted) by E411. A relatively large number of feral cats inhabit the proximities of the Center, facilitating the occurrence of this unusual event. The intruder was treated very gently and showed no signs of abuse from its "foster father." The adult male rhesus held the kitten against his chest most of the time (see Figure 1), but for quadrupedal walking, carried it in his mouth, holding it by the nape of the neck.
The male rhesus kept the young cat in this way until it was removed a day later by the caretakers, to prevent the kitten's death from starvation.
Figure 1: E411 and kitten.
This is the first documented case of interspecies adoption in this Center that involves an adult male. The other case (Martinez & Kessler, 1979) was of an adult female who adopted a live feral rat after the death of her newborn infant.
Reference ;
Martinez, H. S. & Kessler, M. J. (1979). Report of an unusual interspecies adoption by a rhesus monkey. Laboratory Primate Newsletter, 18[3], 6.
-------------------------------------------------------------------
Author's address: P. O. Box 147, Manati, PR 00701.
* * *
Chimpanzee Sperm Bank
Kenneth G. Gould, chief of reproductive biology at Yerkes Regional Primate Center, has established the world's first sperm bank for chimpanzees, and says he has indisputable proof that it works``a female chimp named Alice who is three months pregnant. For more than 3 decades, scientists have known how to freeze human and cattle sperm successfully, but the formulas they used didn't prevent ice crystals from destroying chimpanzee sperm. Dr. Gould began experimenting with various mixtures and freezing rates in an attempt to prevent formation of ice crystals. Eventually he found that chimp sperm had to be frozen more slowly than other species, and also had to be treated with glycerol. Experiments indicate that about 60% of the spermatozoa survive, compared to about a 70% survival rate in cattle sperm banks. Ten females have each been artifically inseminated one time with previously frozen sperm, and one is pregnant and past the time period when spontaneous abortions in chimps are common. A second female may be pregnant.
The new sperm bank will help scientists preserve one of medicine's most valued research animals, which are a threatened species that no longer can be legally captured in the wild. It will help breeders maintain a genetically diverse population of captive chimps in the United States and give scientists a model for developing sperm banks for other primates like gorillas, which are even closer than chimps to extinction in the wild. -- From an article by Hal Straus in The Atlanta Constitution, October 25, 1988.
Lawsuit Against U.C. Dismissed
On August 18, an Alameda County Superior Court judge dismissed a suit brought against the University of California system by Animal Allies, formally the Sacramento branch of People for the Ethical Treatment of Animals. The activists sued to gain access to the university's ten animal care and use committees. The judge ruled that the UC committees are not subject to the state's open meeting law. Therefore, the committees do not have to be open to the general public. Animal Allies plans to appeal the court's decision. -- From NABR Update, 9[19], September 26, 1988.
Pickett Retires
Dr. Betty H. Pickett, Director of the Division of Research Resources (DRR) for the past 6 years, retired on October 1 after 31 years of Federal service. Her NIH career began in 1958 as Executive Secretary of the Behavioral Sciences Study Section in the Division of Research Grants. Later, she was appointed Chief of the Cognition and Learning Section, Division of Extramural Research Programs, NIMH. From 1968 to 1978, she served as Deputy Director of that Division, receiving the DHEW Superior Service Award for her work.
In 1979, Dr. Pickett became Deputy Director, and later Acting Director, of NICHD. In 1982, she was appointed by Dr. James P. Wyngaarden, NIH Director, as Director of the DRR--the component of NIH responsible for creating, developing, and making available a broad array of centers, resources, and institutional support required by the biomedical research activities of the NIH and other research components of the Public Health Service.
Dr. Pickett plans to divide her time between her homes in Washington, DC and Surry, ME. Dr. Robert A. Whitney, Jr., Director of the Division of Research Services, has been named Acting Director of DRR by Dr.Wyngaarden, who plans to amalgamate the two divisions.
ICLAS
The new Governing Board of the International Council for Laboratory Animal Science (ICLAS) and a number of observers met September 17-18 in Liestal, Switzerland. Scholarships were awarded to students from Nigeria, Kenya, and Uganda, and the report of a scholarship holder from Poland was accepted. Secretary General Osmo Hänninen distributed a draft plan for the East African Regional Laboratory Animal Center, which had been prepared on the basis of discussion with Kenyan experts. Past Secretary General Ramsay Hovell reviewed the present phase of work in the preparation of a manuscript for the ICLAS/WHO guidelines for laboratory animal services for developing countries. He was authorized to finalize the editing. It was recommended that a special course on laboratory animal science be organized in Brazil before or after the XXIV World Veterinary Congress, to be held there in 1991. The East African Regional Symposium of Laboratory Animal Science, which will be held in November, 1989, will be the main meeting of ICLAS this year.
NIH Briefs Congressional Aides
The National Institutes of Health (NIH) sponsored a presentation entitled "Why Biomedical Research with Animals is Essential" for congressional aides and staff on June 17, 1988 in Washington, DC. Speakers included Dr. Franklin M. Loew, dean of the Tufts University School of Veterinary Medicine (whose statement is printed in the Fall, 1988, issue of ILAR News) , Dr. William F. Raub, deputy director of NIH, and Dr. Steven A. Rosenberg, chief of the Surgery Branch, National Cancer Institute, NIH.
Costs for New Animal Rules
Regulations on amendments to the Animal Welfare Act of 1985 are expected to be released soon from the Office of Management and Budget, where they have been under review since July. According to the Agriculture Department's Animal and Plant Health Inspection Service (APHIS), the long-awaited "Part 3" of new regulations would cost research institutions about $111 million in capital expenditures on primate facilities.
APHIS's proposed primate rules draw heavily on recommendations from an advisory committee that included ten researchers. They contain a new section on social housing, revise specifications for "primary enclosures," and spell out ways to enrich cage environments. A draft of the proposed rules, described in Science (1988, 242, 662-663) in a "News and Comments" article by Constance Holden, says that primates are "social beings in nature and require contact with other nonhuman primates for their psychological well-being." Therefore, they must be housed with others, in pairs or groups, "whenever possible and consistent with" their health, safety, and well-being.
New prescriptions for cage sizes are set for each of seven categories by weight. They correspond for the most part with recommendations in the NIH "Guide for the Care and Use of Laboratory Animals." The rules state that "the physical environment in primary enclosures must be enriched by providing means of expressing species-typical activities." Special enrichment measures must be designed for juveniles, primates showing signs of psychological distress, and animals used in research that does not involve physical activity.
According to the regulatory impact analysis, a large part of the additional outlay of $111 million will be for new cages. The total number of cages needing replacement is estimated at over 59,000. New recreational equipment is estimated at $100 to $1000 per cage. In addition, 429 facilities are said to need to add building space for primate housing, and 264 to require renovations. How and by whom the costs of all the changes will ultimately be borne remains an unanswered question. The only federal money specifically targeted for animal facilities improvement is a program in NIH's Division of Research Resources. The fiscal 1989 budget for this is $11.6 million, which requires matching funds from the recipient.
* * *
Alexandra C. Bakarich, USAF MEDCEN/SGS, Keesler AFB, MS 39534-5300.
Maj. Alan L. Chedester, AFRIMS, Vet. Med., APO San Francisco, CA 96346-5000.
J. Erwin, Director, Primate Ecology Dept., Diagnon Corp., 11 Taft Court, Rockville, MD 20850.
Susan J. Heath, 5717 Golden Eagle Lane, Roanoke, VA 24018.
Kenneth R. Kaemmerer, Dallas Zoo, 621 E Clarendon, Dallas, TX 75203.
Ernst Knobil, Lab. of Endocrinology, Univ. of Texas Health Science Center, P.O. Box 20708, Houston, TX 77225.
M. Chimes, Abbott Labs D403, 1400 Sheridan Rd., N. Chicago, IL 60064.
Michael Kaplan, Childrens Hospital, Enders Building, 300 Longwood Ave., Boston, MA 02115.
Michale E. Keeling, Univ. Texas System Cancer Center, Environmental Science Park, Rt. 2, Box 151-B1, Bastrop, TX 78602.
Paul Kirk, Schering Corp., P.O. Box 32, Lafayette, NJ 07848.
Steven M. Kuhlman, Dept. Lab. Animal Medicine, TRW Hazleton Lab., 9200 Leesburg Tpk, Vienna, VA 22180.
Scott Line, California Primate Research Center, University of California, Davis, CA 95616.
Dan Lyons, Thomas Jefferson Univ., 644A JAH, 120 Locust St., Philadelphia, PA 19107.
LaVonne Meunier L-70, Smith Kline & French, P.O. Box 1539, King of Prussia, PA 19406-0939.
Richard K. Nakamura, Room 11-105, Parklawn Bldg, 5600 Fishers Lane, Rockville, MD 20857.
Mike Parker, University of Iowa, Animal Care Unit, 400 Medical Laboratories, Iowa City, IA 52242.
Amy Morris-Smith, 1120 E. 38th Place, Tulsa, OK 74105.
Richard Nelson, Lab. Animal Resource Center, Washington State Univ., Pullman, WA 99164-1165.
Arthur W. Rowe, New York Univ. Medical Center, LEMSIP, Long Meadow Rd., Tuxedo, NY 10987.
M. E. Stephens, Dept. of Anthropology, University of Calgary, 2500 University Drive NW, Calgary, Alta. T2N 1N4 Canada. David A. Valerio, Hazleton Res. Products, P.O. Box 7200, Denver, PA 17517.
Janet D. Thomas, 870 East El Camino Real #131, Mountain View, CA 94040.
Pat Ward, American Cyanamid, 56B/236, Middletown Rd., Pearl River, NY 10965.
Vern Weitzel, Dept. of Prehistory & Anthropology, Australian National University, Canberra, ACT 2601 Australia.
* * *
University of Calgary
The Department of Anthropology at the University of Calgary is seeking a PhD with interests in behavioral ecology, paleontology, anatomy and physiology, or any combination of these areas, for an Assistant Professorship in Primatology, appointment effective July 1, 1989. The successful candidate will be expected to develop and maintain a quality research program, publish in refereed journals, and have an established record of excellent teaching.
The department currently offers both BA and BSc programs, as well as a Masters degree. Application for a doctoral program is being actively considered by the department. Normal teaching load is five half-courses per year, exclusive of graduate supervision.
Applications, including a curriculum vitae and names, addresses, and telephone numbers of three referees, should be sent by January 31, 1989 to Dr. Usher Fleising, Head, Department of Anthropology, The University of Calgary, 2500 University Drive N.W., Calgary, Alberta T2N 1N4, Canada.
Salary and rank are commensurate with qualifications and experience. In accordance with Canadian immigration requirements, priority will be given to Canadian citizens and permanent residents of Canada.
University of Texas
A part-time (24 hours per week) behavioral research assistant position is now available at The University of Texas M. D. Anderson Science Park Chimpanzee Breeding Facility. Duties include collecting behavioral data, computer entry of data, and completing preliminary data summary and analysis. A bachelor's degree in a relevant field is required. Annual salary is $10,591. Contact Lydia Elliott or Dr. Mollie Bloomsmith, UTMDACC, Science Park, Veterinary Resources Dept., Route 2, Box 141-B1, Bastrop, TX 78602 [512-321-3991]. The University of Texas M. D. Anderson Cancer Center is an equal opportunity/affirmative action employer.
Wake Forest University
The Department of Comparative Medicine, Bowman Gray School of Medicine, Wake Forest University, seeks a veterinarian at the Assistant or Associate Professor level, in a large, research-oriented academic department. Applicants should have some administrative experience, expertise in medical primatology, and interests in research. The successful candidate will be expected to assume a leadership role for the routine animal health management at the Friedberg Campus (Research Farm), participate in research, and train veterinary residents, postdoctoral fellows, and graduate students. Interested individuals should communicate with Dr. Carolyn F. Moyer, Chairperson, Search Committee, Department of Comparative Medicine, Bowman Gray School of Medicine, 300 S. Hawthorne Road, Winston-Salem, NC 27103 [919-764-3600]. Wake Forest University is an equal opportunity, affirmative action employer.
Primate Foundation of Arizona
A Registrar is wanted, to maintain a computer record system, to account for inventory and health status of a chimpanzee breeding colony, and various other office duties. Some computer experience is desirable, and the applicant must type at least 40 wpm and have good communication skills. Salary is $12,480-13,500, depending upon experience. Excellent benefits. Send cover letter, resume, and references by 15 February 15, 1989, to Jo Fritz, Director, Primate Foundation of Arizona, P.O. Box 86, Tempe, AZ 85280.
* * *
Books
*Nonhuman Primate Studies on Diabetes, Carbohydrate Intolerance,
and Obesity (Monographs in Primatology,
Vol. 12). Charles F. Howard,
Jr. (Ed.). New York: Liss, 1988. 162 pp. [Price:$39.50]
. . Contents: Discrete yet serial indications of the development of
Type 2 diabetes in the rhesus monkey: Marked similarity to human NIDDM,
by N. L. Bodkin & B. C. Hansen. Obesity in pregnancy: Consequences for
maternal glucoregulation and fetal growth, by J. W. Kemnitz, M. J.
Engle, T. J. Flitsch, R. H. Perelman, & P. M. Farrell. Body fat
distribution and atherosclerosis, by C. A. Shively & T. B. Clarkson.
Experimental insulin-deficient diabetes in rhesus monkey: Blood
biochemistry, renal function and structure, and platelet physiology, by
G. Takimoto, C. Jones, A. Bauman, J. Jeffery, E. John, J. Manaligod, &
O. Jonasson. Contributions of the insular amyloidotic islet lesion to
the development of diabetes mellitus in
Macaca nigra, by C. F. Howard, Jr. & T.-Y. Fang. Chronic
hyperinsulinemia and the fetal rhesus monkey: A model of human fetal
macrosomia syndromes, by J. B. Susa. Neonatal hypoglycemia and behavior,
by A. M. Schrier, P. Brady, M. L. Povar, R. Schwartz, & J. B. Susa.
Baboons as a model for research on metabolism, feeding, and the
regulation of body weight, by S. C. Woods, D. P. Figlewicz Lattemann, A.
J. Sipols, & D. Porte, Jr.
*Comparative Behavior of African Monkeys. (Monographs in Primatology,
Vol. 10). Evan L. Zucker,
(Ed.). New York: Liss, 1987. 239 pp. [Price:$49.50]
. . Contents: African monkeys in behavioral research: A 5-year
retrospective analysis, by J. Erwin & E. L. Zucker. Sexual differences
in early social development among captive patas monkeys, by K. M. Loy &
J. Loy. Behavioral development and proximity patterns in captive
DeBrazza's monkeys, by B. C. Kirkevold & C. M. Crockett. Development of
social behavior in vervet monkeys, Sykes' monkeys, and baboons, by C. A.
Bramblett & A. M. Coelho, Jr. Use of space by young African green
(Cercopithecus aethiops sp) and patas (Erythrocebus patas)
monkeys in captivity, by M. R. Clarke, J. R. Kaplan, E. C. Nicks, & A.
R. Grady. Interspecies interactions between free-ranging patas
(Erythrocebus patas) and rhesus (Macaca mulatta) monkeys, by E.
L. Zucker. Dominance and affiliation in the Cercopithecini and
Papionini: A comparative examination, by J. R. Kaplan. Social status and
the distribution of social behavior by adult female patas monkeys: A
comparative perspective, by E. L. Zucker. The sexual behavior of African
monkeys and the question of estrus, by J. Loy. Management and husbandry
of African monkeys in captivity, by M. Bloomstrand & T. L. Maple.
*Ecology and Behavior of Food-Enhanced Primate Groups
(Monographs in Primatology,
Vol. 11). John E. Fa & Charles H. Southwick,
(Eds.). New York: Liss, 1988. 355 pp. [Price:$59.50]
. . Contents: I. Food and Energetics. Feeding behavior of a
food-enhanced troop of Hanuman langurs
(Presbytis entellus) in Jodhpur, India, by P. Winkler. Dynamics of
exploitation: Differential energetic adaptations of two troops of
baboons to recent human contact, by D. L. Forthman Quick & M. W.
Demment. Supplemental food as an extranormal stimulus in Barbary
macaques
(Macaca sylvanus) at Gibraltar -- its impact on activity budgets, by
J. E. Fa. Food and energetics of provisioned wild Japanese macaques
(Macaca fuscata) , by T. Iwamoto. Feeding behavior and activity
patterns of rhesus monkeys
(Macaca mulatta) at Tughlaqabad, India, by I. Malik & C. H.
Southwick. Food habits of rhesus monkeys
(Macaca mulatta) in the north Indian plains, by M. F. Siddiqi & C.
H. Southwick. Time budgets of rhesus monkeys
(Macaca mulatta) in a forest habitat in Nepal and on Cayo Santiago,
by B. M. Marriott.
II. Demography and Life History Patterns. Effects of supplementary
feeding on maturation and fertility in primate groups, by J. Loy.
Dynamics of provisioned and unprovisioned primate populations, by A. M.
Lyles & A. P. Dobson. Life-history patterns of Barbary macaques
(Macaca sylvanus) at Affenberg Salem, by A. Paul & J. Kuester. III.
Behavior and Social Organization. Impact of feeding practices on growth
and behavior of stump-tailed macaques
(Macaca arctoides) , by A. S. Chamove & J. R. Anderson. Relationship
between foraging and affiliative social referencing in primates, by M.
W. Andrews & L. A. Rosenblum. Demography and mother-infant
relationships: Implications for group structure, by C. M. Berman.
Ecological constraints and opportunities: Interactions, relationships,
and social organization of primates, by P. C. Lee. Studies of
food-enhanced primate groups: Current and potential areas of
contribution to primate social ecology, by D. R. Rasmussen.
*Nonhuman Primates in Perinatal Research. Yves W. Brans &
Thomas J. Kuehl (Eds.). New York: Wiley, 1988. 472pp. [Price: $85]
. . A review of research involving the physiology and pathology of
reproduction in nonhuman primates. Contents: I. The Pregnant Primate.
Endocrinology of pregnancy, by E. D. Albrecht & G. J. Pepe.
Pregnancy-induced hypertension: Experimental studies in the nonhuman
primate, with clinical correlation, by P. S. Rao, D. Cavanagh, & R. A.
Knuppel. Anatomy and function of the myometrium, by R. H. Hayashi.
Perspective on the pregnant primate, by R. F. Williams & K. Adamsons.
II. The Embryo. Fertilization and preimplantation embryonic development,
by W. R. Dukelow & M. T. Ridha. Implantation and early embryonic
development in primates, by A. C. Enders & A. G. Hendrickx. In vitro
development of the primate embryo, by V. Pope, E. Pope, & L. Beck.
Placental morphology, by M. L. Houston. Perspective on the embryo, by
D. C. Kraemer. III. The Fetus. Bioelectronic
monitoring of the fetus, by R. H. Hayashi. Amniotic fluid volume,
composition, ingestion, and digestion by the fetus, by Y. W. Brams.
Model of diabetes: Fetal hyperinsulinemia and macrosomia, by J. B. Susa
& R. Schwartz. Chronic instrumentation of the fetal primate, by Y.
Murata, S. Fujisaki, C. Endo, & C. B. Martin, Jr. Intrauterine
intervention: Fetal surgery in the nonhuman primate, by A. W. Flake, M.
R. Harrison, & N. S. Adzick. Model of cigarette smoking, by W. R. Rogers
& T. J. Kuehl. Perspective on the fetus, by C. B. Martin, Jr. & Y.
Murata. IV. The Neonate. A. Pulmonary and Cardiovascular Functions.
Pathology of perinatal lung disease, by J. J. Coalson. Surfactant
studies, by R. V. Kotas. Cardiopulmonary studies and high-frequency
ventilation, by R. A. deLemos, A. Guajardo, & K. Meredith. Structure and
function of the developing diaphragm, by R. J. M. McCarter & L. C.
Maxwell. Baboon model for meconium aspiration, by J. C. Carey. The
development of extracorporeal membrane oxygenation for the treatment of
human neonates using nonhuman primates as models, by J. D. Cornish & D.
R. Gerstmann. Pharmacology of cerebral and mesenteric arteries, by S.
Hayashi & M. K. Park. Perspective on neonatal pulmonary cardiovascular
functions, by R. V. Kotas. B. Body Composition, Nutrition, Behavior, and
Infections. Fetal and neonatal body composition, by Y. W. Brans.
Bilirubin metabolism in fetus and neonate, by C. E. Cornelius.
Behavioral testing, by W. R. Rogers. A rhesus monkey model of neonatal
group B streptococcal infection, by V. G. Hemming & W. T. London. Fetal,
neonatal, and maternal growth and hydration: A brief overview, by D. B.
Cheek, O. M. Petrucco, & A. Gillespie. Conclusion, by Y. W. Brans & T.
J. Kuehl.
*Primate Adaptation and Evolution. John G. Fleagle. New York:
Academic Press, 1988. 486 pp. [Price: $39.95]
. . This was written as a textbook for advanced undergraduate and
beginning graduate students in anthropology and biology. It covers
the comparative anatomy, behavioral ecology, and paleontology of the
order Primates by presenting the major groups of living and extinct
primates as a series of adaptive radiations.
*Comparative Anatomy of the Cerebral Cortex of Man and Monkey. A. A. Khachaturian. Moscow: Izdatel'stvo "Nauka," 1988. 215 pp. [In Russian. Price: 3 Rubles]
Bibliographies
*Behavioral Observations of Feral and Free-ranging Cebidae: A Bibliography, 1980-1988. J. B. Williams. Seattle: Primate Information Center, 1988. (221 citations, species & subject indexes) [Price: $6.50. Send order to Primate Information Center, Regional Primate Research Ctr., SF-50, Univ. of Washington, Seattle, WA 98195.]
*Behavior of Captive Cynomolgus Monkeys (Macaca fascicularis): A Bibliography. J. B. Williams. Seattle: Primate Information Center, 1988. (172 citations) [Price: $6.50. Ordering information same as above]
*Adult Male-Infant Interaction: A Bibliography, 1981-1988. J. B. Williams. Seattle: Primate Information Center, 1988. (217 citations, primate index) [Price: $6.50. Ordering information same as above]
*Physiological and Pharmacological Correlates of Dominance: A Bibliography, 1973-1988. J. B. Williams. Seattle: Primate Information Center, 1988. (196 citations, primate & subject indexes) [Price: $6.50. Ordering information same as above]
*Primate Names: Scientific and Vernacular -- Discussions, Claims, Controversies, Revisions, Histories: A Bibliography, 1978-1988. J. L. Pritchard. Seattle: Primate Information Center, 1988. (140 citations, primate index) [Price: $6.50. Ordering information same as above]
*Group Behavioral Variations in Feral and Free-Ranging Nonhuman Primates: A Bibliography, 1978-1988. J. B. Williams. Seattle: Primate Information Center, 1988. (216 citations, primate index) [Price: $6.50. Ordering information same as above]
*Infectious Diseases (Bacterial, Viral, Mycotic, and Endoparasitic) of Feral Macaques (Macaca) and Baboons (Papio): A Bibliography, 1972-1988 with selected earlier citations. J. L. Pritchard. Seattle: Primate Information Center, 1988. (156 citations, primate & subject indexes) [Price: $6.50. Ordering information same as above]
*Behavioral Observations of Feral and Free-Ranging Baboons (Papio and Theropithecus): A Bibliography, 1980-1988. J. B. Williams. Seattle: Primate Information Center, 1988. (228 citations, species index) [Price: $6.50. Ordering information same as above]
Proceedings
*Laboratory Animal Studies in the Quest of Health and Knowledge.
H. A. Rothschild, A. Rosenkranz, & F. A. Moura Duarte (Eds.). 1987.
331 pp. [Price:
$10 plus $2 postage and handling. Order from H. A. Rothschild,
INFAR-EPM, Rua Tres de Maio, 100, 04044 São Paulo, SP Brazil]
. . Proceedings of a scientific meeting held in Aguas de Lindóia, SP,
Brazil, November 18-21, 1986, under the sponsorship of the
Multi-Institutional Center for Laboratory Animal Science, the Federation
of the Societies for Experimental Biology, and the International Council
for Laboratory Animal Science. Besides papers and lectures on nutrition,
ethics and legislation, teaching and training, alternatives, housing and
environment, and sanitary monitoring for laboratory animals in general,
there was a session on neotropical primates as models for biomedical
research and for production and control of medicines.
Reports
*Use of Laboratory Animals in Biomedical and Behavioral Research.
National Research Council. Washington, DC: National Academy Press, 1988.
102 pp.
[Price: $9.95 in U.S., Canada, & Mexico; $11.95 elsewhere. Order from
National Academy Press, 2101 Constitution Ave. NW, Washington DC 20418]
. . A statement on the costs and benefits, to humans and animals,
of using animal subjects, with recommendations for the future. Two
members of the committee, one of whom did not sign the report, included
individual statements. In addition, one other member, John Kaplan, while
in agreement with the report, published a
Perspective article in Science ( 1988, 242, 839-340), which
emphasized the political dimension of the issue of animal rights.
*Biosafety in Microbiological and Biomedical Laboratories (Second Edition). Centers for Disease Control & the National Institutes of Health. Washington, DC: U.S.G.P.O., 1988. [Single copies are free to academic institutions and private laboratories from NIH/DS, 31-1C02, Bethesda, MD 20892. Copies for other institutions or multiple copies are $3.75 per copy (stock no. 107-040-000508-3) from the Superintendent of Documents, U.S. Government Printing Office, Washington, DC 20402]
Special Journal Issues
*A Fifteen Year Retrospective on the Endangered Species Act. Endangered Species Update. August, 1988, 5[10].
Animal Models
*Naturally occurring chronic gastritis and
C pylori infection in the Rhesus monkey: A potential model for
gastritis in man. Baskerville, A. & Newell, D. G. (Public Health
Laboratory Service, Centre for Applied Microbiology and Research, Porton
Down, Salisbury, Wiltshire SP4 0JG, England).
Gut, 1988, 29, 465-472.
. . Histological examination of the stomachs of 11 rhesus monkeys at
autopsy showed chronic gastritis in a high proportion of all ages.
Gastric Campylobacter-like organisms apparently identical to human
C. pylori were cultured and/or detected immunohistologically in
several animals. Two of 5 cynomolgus monkeys examined had
gastritis, which may be associated with the presence of
C. pylori. Ten baboons examined
did not have gastritis, nor was C. pylori
cultured from their stomachs.
Animal Welfare
*Experts ponder simian well-being. Holden, C.
Science, 1988, 241, 1753-1755.
. . A report on the conference on psychological well-being of captive
primates held in Boston this September, under the sponsorship of Tufts
University, the New England Primate Center, and the University of
Massachusetts, with a grant from the American Psychological Association.
Behavior
*Social factors influencing choice of food source in a captive group
of chimpanzees
(Pan troglodytes) . Gust, D. A. (Yerkes Regional Primate Research
Center Field Station, Emory Univ., 2409 Taylor Rd., Lawrenceville, GA
30243).
American Journal of Primatology, 1988, 16, 81-87.
. . A group of 16 chimpanzees maintained in a large, outdoor compound
chose among three feeding stations that were spatially separated and
located along the periphery of the compound. The study provides
quantitative data supporting postulations that factors such as social
relationship, kinship, and estrous cycle stage affect the composition of
groupings of chimpanzees at feeding stations, and the results are
consistent with field observations.
*Preliminary report on the activity cycle and diet of the
red-bellied lemur
(Lemur rubriventer) in Madagascar. Overdorff, D. (Duke Univ.
Primate Center, 3705 Erwin Rd., Durham, NC 27705).
American Journal of Primatology, 1988, 16, 143-153.
. . Data were collected on diet, patterns of activity, and group
composition of red-bellied lemurs in a southeastern rainforest of
Madagascar during June and July 1986. In June, the group's major food
resource was fruit, and they maintained a strict diurnal activity cycle.
In July, they fed on flowers (80%), fruits (10%), and leaves (10%), and
were nocturnal as well as diurnal. The transition from a diurnal to a
random, 24-hour activity pattern may be an adaptive strategy to avoid
predation by diurnal raptors.
*Sexually dimorphic transitions revealed in the relationships of
yearling rhesus monkeys following the birth of siblings. Holman, S. D. &
Goy, R. W. (MRC Neuroendocrine Development & Behaviour Group, AFRC Inst.
of Animal Physiology, Babraham, Cambridge, CB2 4AT, U.K.).
International Journal of Primatology, 1988, 9, 113-133.
. . Social interactions of 1-year-old animals 6 weeks before the birth
of siblings were compared with interactions occurring 6 weeks after.
Among yearling females, no dramatic changes in interactions with their
mothers or with other group members were detected, but yearling males
refocused their attentions away from their mothers and toward other,
often unrelated members of the group.
*Maternal time budgets of gelada baboons. Dunbar, R. I. M. & Dunbar,
P. (Dept. of Anthropology, University College London, Gower St., London
WC1E 6BT, U.K.).
Animal Behaviour, 1988, 36, 970-980.
. . Data from gelada baboons,
Theropithecus gelada, were used to evaluate J. Altmann's model of
maternal time budgets. The results provide substantial confirmation of
the model's general principles: the time that a female devoted to
feeding increased with the age of her infant roughly in line with the
model's predictions. The results suggest that Altmann's assumption that
the additional demand for feeding time is taken out of social time is
only partially correct. Social time was conserved as far as
possible, with extra feeding time being taken out of resting time until
much of the available resting time was used up. Then the female released
some of her social time, but still sought to preserve those social
relationships that are most important to her long-term reproductive
interests.
*Resolution of aggressive conflicts by immature long-tailed macaques
Macaca fascicularis. Cords, M. (Ethologie und Wildforschung,
Zoologisches Inst., Univ. Zürich-Irchel, Winterthurerstrasse 190, 9057
Zürich, Switzerland).
Animal Behaviour, 1988, 36, 1124-1135.
. . The behavior following aggressive interactions differed from that
following friendly or neutral control interactions among unrelated
immature male
M. fascicularis. The behavior characterizing encounters between
former opponents differed in the two situations, with more affiliative
interactions post-conflict. The pattern of interactions between former
opponents is interpreted as a repair mechanism for relationships
strained by aggression.
*Experimental matrilineal inheritance of rank in female Japanese
macaques. Chapais, B. (Dépt. d'Anthropologie, Univ. Montréal, C.P.
6128, Montréal, PQ H3C 3J7, Canada).
Animal Behaviour, 1988, 36, 1025-1037.
. . In a group of 17 Japanese macaques comprised of three families
with similar age-sex compositions, experimental female subgroups were
formed such that in each of them a female (dependent) was given more
alliance power than a dominant target. Results indicate that in each of
the subgroups that was tested the dependent female inherited the rank of
her ally (mother or older sister) above same-age or older targets. The
fact that females who were given more alliance power did not solicit
their ally, or challenge the target females, before they were aided by
their ally suggests that aggressive interventions are the primary
mechanism of rank inheritance. The role and dynamics of the recognition
of alliances in the establishment of rank relationships are discussed.
*Experimental rank reversals among peers in
Macaca fuscata: Rank is maintained after the removal of kin
support. Chapais, B. & LaRose, F. (Address same as above).
American Journal of Primatology, 1988, 16, 31-42.
. . Experimental subgroups were formed by placing together peers from
different families and a single kin (ally) of one subordinate peer
(mother or older sister). These experiments induced a series of rank
reversals whereby the subordinate peer, whose ally was present,
outranked the dominant peers. Following the rank reversals, the ally of
the newly dominant peer was removed and the peers left together. All
five experimentally inverted rank orders of peers remained stable.
*Individual and group effects on early caregiver-infant
relationships in red-bellied tamarin monkeys. Pryce, C. R. (Dept. of
Anthropology, University College London, Gower St., London WC1E 6BT,
U.K.).
Animal Behaviour, 1988, 36, 1455-1464.
. . The relationship between changes in caregiver-caregiver and
caregiver-infant behavior was studied during the infants' first month in
10 family groups of red-bellied tamarins with twin infants. Each
caregiver initiated carrying by retrieving an infant from another
caregiver, and sometimes retained an infant against a retrieve attempt
by another caregiver in order to continue carrying it. Results indicate
that competition to care for infants has a direct effect on the early
development of each caregiver-infant relationship within the family
group.
*Mating season effects on mother-infant conflict in Japanese
macaques,
Macaca fuscata. Worlein, J. M., Eaton, G. Gray, Johnson, D. F., &
Glick, B. B. (G. G. Eaton, Dept. of Reproductive Biology & Behavior,
Oregon Regional Primate Research Center, 505 N.W. 185th Ave., Beaverton,
OR 97006).
Animal Behaviour, 1988, 36, 1472-1481.
. . The impact of seasonal mating on the mother-infant relationship
during the first year of the infant's life was documented in a confined
troop of Japanese macaques. During the weeks in which the mothers were
being mounted, negative behavior directed towards the infants increased.
As maternal rejections and punishments escalated, the infants displayed
regressive behavior. The mother-infant relationship undergoes a period
of conflict during the mating season not because the mother is weaning
her infant, but because it is to her reproductive advantage to regulate
the times in which the infant is allowed to suckle.
*Arbeitsteiliges und kooperatives Problemlöseverhalten in
Primatengruppen [Labor-division and co-operation in problem-solving
primate groups]. Vössing, A. (Zoologisches Inst. der Univ. Münster,
Abt. für Verhaltensforschung).
Zoologische Beiträge, 1987, N. F. 31, 305-340.
. . A group of vervets (4 male, 7 female) and a group of lemurs (2
male, 4 female) were daily given a box, containing food and fastened
with a variety of latches. One vervet opened the latches and box in 95%
of the cases, while 2 lemurs shared 75% of the solutions.
*Changes in the size, composition, and reproductive condition of
wild marmoset groups
(Callithrix jacchus jacchus) in north east Brazil. Scanlon, C. E.,
Chalmers, N. R., & Monteiro da Cruz, M. A. O. (Biology Dept., The Open
University, Milton Keynes, MK7 6AA, England).
Primates, 1988, 29, 295-305.
. . Five groups of
Callithrix jacchus jacchus were studied in the field. Group size
and composition were similar to that described for other callitrichids,
but changes in group membership occurred much more frequently than
previously reported. It is argued that marmoset social organization is
best studied at the level of the whole population within an area, rather
than at the level of the individual group.
*Responsiveness to objects in two social groups of tufted capuchin
monkeys
(Cebus apella). Visalberghi, E. (Ist. di Psicologia, CNR, Via
Aldrovandi, 16b, Roma 00197 Italy).
American Journal of Primatology, 1988, 15, 349-360.
. . The responses of two social groups of
Cebus apella to novel or familiar objects were examined, both in
scarce and abundant conditions. The aim was to obtain a description of
capuchins' general norms of behavior toward objects, focusing on the
variability across sex/age classes. Sex and age significantly affected
the behavioral pattern of interaction with blocks. Dominance did not
seem to constrain monkeys' responsiveness.
*Long-term retention of self-recognition by chimpanzees. Calhoun, S.
& Thompson, R. L. (R. L. Thompson, Dept. of Psychology, Hunter College,
CUNY, New York, NY 10021).
American Journal of Primatology, 1988, 15, 361-365.
. . Two young chimpanzees showed retention of self-recognition after 1
year without access to mirrors. The animals had been positive on the
Gallup dye test. One year later they were once again anesthetized,
marked on the brow and ears, and following recovery, exposed
individually to a large mirror. Both demonstrated mirror-oriented
mark-directed responses. Time-sample observations of behavior toward the
mirror further support self-recognition.
*Inferring patterns of aggression from red howler monkey injuries.
Crockett, C. M. & Pope, T. (13034 1st Ave. NE, Seattle, WA 98125-3005).
American Journal of Primatology, 1988, 15, 289-308.
. . Thirty-eight percent of 119 howler monkeys, examined at capture,
showed scars or other evidence of "damage." Sociobiological costs
of aggression in red howlers are difficult to assess because many
howlers, despite frequent and severe injury, subsequently survive
and reproduce. The authors recommend caution in inferring mortality
from injuries, especially when social mobility and emigration out of
the study area are common.
*Orangutan diet, range, and activity at Tanjung Puting, Central
Borneo. Galdikas, B. M. F. (Orangutan Research and Conservation Project,
Tromol Pos 1, Pangkalan Bun, Kalimantan Tengah, Indonesia).
International Journal of Primatology, 1988, 9, 1-35.
. . This study focuses on an orangutan population in a swampy lowland
area near sea level. The animals were predominantly frugivorous, but the
overall variety in their diet was remarkable; 317 different food types
have been identified, including fungus, insects, and honey. Contact with
other orangutans usually increased a focal animal's day length, day
range, and amount of time spent moving. This suggests that foraging
alone maximized each orangutan's foraging returns by minimizing the day
range traveled. Orangutan solitariness is the result of large body size
and of a predominantly frugivorous and opportunistic diet.
*The variable social organization of Hanuman langurs
(Presbytis entellus) , infanticide, and the monopolization of
females. Newton, P. N. (Animal Ecology Research Group, Dept. of Zoology,
Oxford OX1 3PS, England).
International Journal of Primatology, 1988, 9, 59-77.
. . Data from 24 wild populations of Hanuman langurs are used to test
hypotheses seeking to explain variation in troop structure and the
incidence of infanticide. The occurence of infanticide is associated
with a one-male troop structure and not with a high density. It is
suggested that a one-male troop structure will arise if a male is able
to monopolize a group of females, a multimale troop if he cannot.
*A case of offspring desertion by a female chimpanzee and the
behavioral changes of the abandoned offspring. Hiraiwa-Hasegawa, M. &
Hasegawa, T. (Dept. of Anthropology, Faculty of Science, Univ. of Tokyo,
Hongo, Bunkyo-ku, Tokyo, 113 Japan).
Primates, 1988, 29, 319-330.
. . This is the first record of an immature chimpanzee (a
five-year-old male) abandoned by its emigrating mother, although a
similar phenomenon has been reported for gorillas and Hanuman langurs.
The abandoned offspring followed an adult male of the group, and that
male gave most care to it. The youngster's time spent on grooming others
increased, the time spent on being groomed decreased, and the rate of
aggression toward him increased after his mother's desertion.
Care
*Psychological well-being of primates in captivity. Novak, M. A. &
Suomi, S. J. (Dept. of Psychology, Univ. of Massachusetts, Amherst, MA
01003).
American Psychologist, 1988, 43, 765-773.
. . No consensus presently exists in terms of how to define or
identify psychological well-being in primate subjects. A strategy is
proposed for defining psychological well-being that includes assessment
of physical health, comparison with species-normative behavioral
repetoires, detection of distress, and evaluation of coping responses.
This set of definitions is then used to characterize prototypical
primate laboratory environments in terms of fostering psychological
well-being. The importance of factors other than housing, such as
species-specific characteristics, rearing histories, and phenotypic
differences, is also emphasized. It seems unlikely that simple
prescriptions will be broadly applicable across the whole range of
captive primates.
*Developing effective social and environment enrichment strategies
for macaques in captive groups. O'Neill, P. (NIH Animal Center, 22766
Club Hollow Rd., Dickerson, MD 20842).
Lab Animal, 1988, 17[4], 23-24, 27-28, 31-36.
. . A group of 17 to 20 rhesus monkeys representing 3 matrilines are
free-ranging in a 5-acre enclosure for 8 months in the year, then housed
indoors in an enriched playroom interior for the winter. Data was
collected to determine what types of activity 4 different age groups
engaged in during a 6-week period in winter. The data show both striking
differences and similarities among the age groups. Toys and apparatus
introduced into the cage environment are not simply alternatives taking
the place of social interactions, but rather are tools for redirecting
and in some cases enhancing or stimulating social interactions primarily
through play.
*Rearing cynomolgus monkeys
(Macaca fascicularis) on surrogate mothers with bottle feeding.
Timmermans, P. J. A., Röder, E. L., & Kemps, A. M. L. J. (Dept. of
Comparative & Physiological Psychology, Univ. of Nijmegen, P. O. Box
9104, 6500 HE Nijmegen, the Netherlands).
Laboratory Animals, 1988, 22, 229-234.
. . Fifty-two cynomolgus monkeys were reared in peer groups on
surrogate mothers and fed by bottle during the first months of life. A
description of the technique of bottle feeding and the structure of
surrogate mothers is presented, along with a growth curve. The weight of
mother-reared infants stayed within the standard deviation of the mean
weight of the artificially-reared subjects.
*The social behaviour of hand-reared siamang at Adelaide Zoo.
Parker, S., St. Clair, M., & Wheeler, L. (Reprint requests to D.
Langdon, Adelaide Zoo, Frome Rd., Adelaide, SA 5000, Australia).
Australian Primatology, 1988, 3[2/3], 3-5.
. . Description of an unsuccessful attempt to introduce two male
infant siamangs to a foster mother.
*Food distribution, dominance, and aggressive behaviors in bonnet
macaques. Boccia, M. L., Laudenslager, M., & Reite, M. (Dept. of
Psychiatry, C-268, Univ. of Colorado Health Sciences Center, 4200 East
Ninth Ave., Denver, CO 80262).
American Journal of Primatology, 1988, 16, 123-130.
. . This study examines the impact of manipulation of the distribution
of food resources in a laboratory-housed group of bonnet macaques to
assess the impact of distribution on aggressive behaviors. Food was
restricted to a space which allowed access to only one or two
individuals at a time or was dispersed throughout the living enclosure
to allow universal access in alternating periods. During periods of
clumped distribution, the monkeys obtained access to the food in order
of their dominance status. In addition, overall levels of aggressive
behaviors increased, submissive behavior increased, and play decreased
during those periods. These results confirm the findings from field
studies indicating increases in agonism with restriction of food
resources.
*Formula-fed and breast-fed baboons: Weight growth from birth to
adulthood. Glassman, D. M. & Coelho, A. M., Jr. (Dept. of Sociology &
Anthropology, Southwest Texas State Univ., San Marcos, TX 78666).
American Journal of Primatology, 1988, 16, 131-142.
. . Longitudinal weight data were collected from two groups, each with
11 females and 11 males, of savannah baboons
(Papio cynocephalus anubis) for the total period of growth from
birth to approximately 8 years of age. One group were
fed a standard infant formula by a nursery care technician for the first
18 weeks of postnatal life, while the other
was breast-fed by their mothers for an equivalent period of time. After
18 weeks, the animals were removed from either nursery or mother and
assigned to one of four age-cohort social groups, each balanced for
treatment and gender. No significant weight differences were found
between rearing conditions for the initial 15 weeks of growth.
However, breast-fed females averaged greater weight at all later ages,
and had a slightly more intense adolescent growth spurt than their
bottle-fed peers.
*Successful feeding enrichment for captive chimpanzees. Bloomsmith,
M. A., Alford, P. L., & Maple, T. L. (Univ. of Texas System Cancer
Center, Science Park Veterinary Resources Dept., Rt. 2, Box 151-B1,
Bastrop, TX 78602).
American Journal of Primatology, 1988, 16, 155-164.
. . Four feeding enrichment procedures were implemented simultaneously
on 41 adult chimpanzees in six social groups in the experimental phase
of this ABA design study. The procedures were planned to elicit
time-consuming, noncompetitive feeding. 270 hours of data were collected
over a 6-month period. The barrage of feeding enrichment techniques
significantly reduced agonism and abnormal behaviors.
Conservation
*Madagascar's lemurs: On the edge of survival. Jolly, A.
National Geographic, 1988, 174, 132-161.
. . A report, with photographs by Frans Lanting, on conservation
efforts on Madagascar.
*Experimental technique for the conservation of private lands.
Horwich, R. H. & Lyon, J. (RD 1, Box 96, Gay Mills, WI 54631).
Journal of Medical Primatology, 1988, 17, 169-176.
. . A community-based sanctuary for black howler monkeys
(Alouatta pigra) has been established on 18 square miles of
riverine forest along the Belize River, Belize. It is based on
voluntarily signed pledges by subsistence farmers to abide by management
plans for their lands. Conservation-education programs which impact
favorably on the villagers are being carried out.
*Rain Forest Conservation: A Plea for a More Dynamic, D.I.Y.
Approach. W. R. Kingston. [Available from W. R. Kingston, The
Old Smithy, Bishops Frome, Worcester WR6 5BA, U.K.]
. . A proposal for carefully controlled commercial and tourist
facilities in threatened tropical rain forests, which would
simultaneously protect and make self-supporting endangered habitats and
species.
Disease
*Reversal of chloroquine resistance in malaria parasite
Plasmodium falciparum by desipramine. Bitonti, A., Sjoerdsma, A.,
McCann, P. P., Kyle, D. E., Oduola, A. M. J., Rossan, R. N., Milhous, W.
K., & Kavidson, D. E. Jr. (Merrell Dow Research Inst., Cincinnati, OH
45215).
Science, 1988, 242, 1301-1303.
. . Desipramine was found to be one of
the most effective compounds yet described for the reversal of
chloroquine resistance both in vitro and in vivo.
*Relearning the lessons of Herpes B virus: A report for the monkey
handler. Cummins, L. B. (White Sands Research Center, 2512 Christina
Pl., Alamogordo, NM 88310).
Lab Animal, 1988, 17[3], 25-27.
. . Two basic questions are asked: What have we learned from the 1987
Herpes B virus outbreak, and what must we do differently in the future
to prevent this rare but serious disease? The most important factor in
attending to an exposure is having a procedure in place well in advance
of the time it is needed. Until new developments are introduced to
reduce exposure to Herpes B virus and to control infections when they
occur, we are left with the preventive measures we have relied on for so
long``but we must apply them diligently and not become negligent.
*Spontaneous and experimental hepatitis A in Old World monkeys.
Shevtsova, Z. V., Lapin, B. A., Doroshenko, N. V., Krilova, R. I.,
Korzaja, L. I., Lomovskaya, I. B., Dzhelieva, Z. N., Zairov, G. K.,
Stakhanova, V. M., Belova, E. G., & Sazhchenko, L. A. (USSR Academy of
Medical Sciences, Inst. of Experimental Pathology & Therapy, P.O. Box
66, Gora Tapetziya, Sukhumi 384900, USSR).
Journal of Medical Primatology, 1988, 17, 177-194.
. . Virologic, serologic, biochemical, and morphological data
characterizing spontaneous hepatitis A (HA) in cynomolgus macaques and
green monkeys are reported. Experimental HA was induced in macaques as a
result of infection with human hepatitis A virus (HAV-h). Disease
similar to human HA was induced in cynomolgus macaques by HAV isolates
from spontaneously sick rhesus and green monkeys. This experimental
model of HA in macaques can be used for vaccine and anti-viral
preparation testing.
*Use of BCG vaccines in the control of tuberculosis: A joint
statement by the ACIP and the Advisory Committee for Elimination
of Tuberculosis.
Morbidity and Mortality Weekly Report, 1988, 37, 663-664, 669-675.
. . Immunization with BCG vaccine lowers the risk of serious
complications of primary TB in children. However,
it is no longer recommended for health
care workers or other adults at high risk for acquiring TB infection. In
addition, BCG should not be given to perons who are immunocompromised,
including those with human immunodeficiency virus infection.
*Cholera vaccine: Recommendations of the Immunization Practices Advisory Committee. Morbidity and Mortality Weekly Report, 1988, 37, 617-618, 623-624.
*Simian AIDS ELISA: Sensitivity, specificity and predictive values
based on a comparison with Western blot technique. Spinner, A. &
Sutjipto, S. (California Primate Research Center, Univ. of California,
Davis, CA 95616).
Laboratory Animal Science, 1988, 38, 568-572.
. . One hundred thirty-one rhesus macaques were tested for the
presence of antibody against SAIDS type D retrovirus by both ELISA and
Western blot techniques. Sensitivity, specificity, and predictive values
for the SAIDS ELISA were calculated based on a
comparison with Western blot results. Sera was tested from two distinct
populations, endemic (71 macaques from a half-acre outdoor corral where
SAIDS was first recognized) and control (60 rhesus macaques from both
indoor and outdoor areas where the disease was not recognized). ELISA is
shown to be a useful screening tool based on its high sensitivity for
both populations, providing a rapid and economical way to diagnose and
manage SAIDS in captive nonhuman primate colonies.
*Hepatocystis in populations of baboons (Papio hamadryas
s.l.) of Tanzania and Ethiopia. Phillips-Conroy, J. E., Lambrecht, F.
L., & Jolly, C. J. (Dept. of Anatomy and Neurobiology, Washington Univ.,
St. Louis, MO 63110).
Journal of Medical Primatology, 1988, 17, 145-152.
. . Bloodsmears from 119 yellow baboons
(Papio hamadryas cynocephalus) in 6 groups in Mikumi National Park,
Tanzania, and from 189 olive
(P. h. anubis) , hamadryas (P. h. hamadryas) , and hybrid baboons
in three groups in Awash National Park, Ethiopia, showed differing
levels of infection with
Hepatocystis simiae. 0% to 42% in the Tanzanian groups were
carriers of the parasite, but none of the Ethiopian baboons was
positive. The remarkable absence of
H. simiae in the Ethiopian baboons is believed to be related to
environmental factors that exclude transmission in the absence of the
Culicoides vector.
Genetics
*Application of DNA fingerprinting probes to Old World monkeys.
Weiss, M. L., Wilson, V., Chan, C., Turner, T., & Jeffreys, A. J. (Dept.
of Anthropology, 139 Manoogian Hall, Wayne State Univ., Detroit, MI
48202).
American Journal of Primatology, 1988, 16, 73-79.
. . Analysis of tandem-repetitive minisatellite DNA in humans provides
great discriminatory power in paternity identification and
individualization of tissue samples. The molecular probes used to study
these hypervariable loci are here applied to DNA isolated from related
animals in four species of Old World monkeys. Probes 33.6 and 33.15 are
shown to cross-hybridize with variable minisatellites in
Macaca fuscata, M. silenus, Erythrocebus patas, and
Colobus guereza. The DNA fingerprint patterns are consistent with
germ-line stability and mendelian inheritance. Application of DNA
fingerprinting to Old World monkeys thus provides a new tool for
identifying parentage and can thereby aid in colony management. The
technique can also help to elucidate the role of behavioral factors in
evolution. The breadth of applicability in other primates remains to be
mapped.
Instruments and Techniques
*Real-time ultrasonic evaluation of the baboon pregnancy: Biometric
measurements. Farine, D., MacCarter, G. D., Timor-Tritch, I. E., Yeh,
M.-N., & Stark, R. I. (R. I. Stark, Dept. of Pediatrics, Babies
Hospital, 622 W. 168th St., New York, NY 10032).
Journal of Medical Primatology, 1988, 17, 215-221.
. . Ultrasonic and direct biometric measurements of nine fetal baboons
were made during 23 evaluations. Both the femur length and the
biparietal diameter were correlated to gestational age. These initial
results have facilitated intrauterine surgery. The data presented will
help in determining adequate fetal growth and in dating baboon
pregnancies.
*Comparison of blood flow parameters measured by Doppler ultrasound
and radionuclide techniques in the baboon
(Papio ursinus) . Bosman, H., Dormehl, I. C., Hordijk, C.,
Maree, M., & Hugo, N. (I. C. Dormehl, AEC Inst. of Life Sciences,
Faculty of Medicine, Univ. of Pretoria, P.O. Box 2034, Pretoria 0001,
South Africa).
Journal of Medical Primatology, 1988, 17, 223-231.
. . Ascending aortic blood velocity was measured in the baboon by
using continuous wave Doppler ultrasound. The blood flow parameters thus
obtained were compared to those by the standardized radionuclide
technique. It appears that, due to the anatomical position of the
ascending aorta and brachiocephalic trunks in relation to the ultrasound
beam, Doppler ultrasound does not provide an accurate method of
measuring aortic blood velocity in the baboon.
*A backpack system for chronically instrumented unrestrained
baboons. Maclean, J. M., Phippard, A. F., Gillin, A. G., & Horvath, J.
S. (Renal Hypertension Unit, Royal Prince Alfred Hospital, Camperdown,
NSW 2050, Australia).
Australian Primatology, 1988, 3[2/3], 7-8.
. . A system to protect and immobilize exteriorized instruments in
conscious unrestrained baboons, while allowing periodical access for
experimental work. The backpack has a removable transparent cover, which
allows remote inspection of the catheters and their exit sites between
the scapulae. A second backpack cover attaches to a flexible stainless
steel tether, which carries catheter extension tubing or electrical
connections to recording equipment during experimental sessions. A plate
located inside the backpack between the base and cover provides a
mounting point within the backpack on which catheter ends and electrical
connections can be fixed.
*Spider monkey home ranges: A comparison of radio telemetry and direct
observation. Fedigan, L. M., Fedigan, L., Chapman, C., & Glander, K. E.
(Dept. of Anthropology, Univ. of Alberta, Edmonton, Alberta, Canada).
American Journal of Primatology, 1988, 16, 19-29.
. . The ranging patterns of 2 male and 5 female spider monkeys were
studied with the use of radio telemetry in Santa Rosa National Park,
Costa Rica. The probability of encountering a radio-collared spider
monkey in a 3-hour search using radio telemetry (0.91) was much greater
than using a visual search (0.20), and telemetric data resulted in a
larger estimate of mean home range size than did observational data,
when all subjects were compared. However, the difference appeared to be
owing to the presence of male ranges in the telemetric, but not the
observational, data. When comparison was made between radio-tracking and
observation results for range size for adult females only, there was no
significant difference.
Pharmacology & Anesthesia
*Alphaxolone-alphadolone for anesthesia of squirrel monkeys of
different ages. Lögdberg, B. (Inst. of Environmental Health, Univ. of
Lund, Sölvegatan 21, S-223 62 Lund, Sweden).
Journal of Medical Primatology, 1988, 17, 163-167.
. . This report describes 75 cases of reliable 1-2 hour surgical
anesthesia of newborn to adult squirrel monkeys, with rapid induction
and recovery, using a single intramuscular dose of
alphaxolone-alphadolone (doses between 11.5 and 30 mg/kg body weight).
*Obstetric analgesia and infant outcome in monkeys: Neonatal
measures after intrapartum exposure to meperidine or alfentanil. Golub,
M. S., Eisele, J. H., & Donald, J. M. (California Primate Research
Center, University of California, Davis, CA 95616).
American Journal of Obstetrical Gynecology, 1988, 158, 1219-1225.
. . Neurobehavioral evaluations were conducted in neonatal monkeys
(Macaca mulatta) exposed to narcotic analgesics during labor
through their dams. They were compared to controls whose dams received
no analgesic. Drug-exposed infants showed neurobehavioral effects over
the first 3 days of life. Drug exposure was also associated with
increased elicited muscle tone early in the neonatal period and earlier
maturation of sitting, standing, and walking, as well as lower
hematocrits and an earlier onset of linear growth rate as reflected in
body weight. No qualitative or quantitative differences were noted
between the two drug treatments.
Physiology
*A study of postnatal appendicular skeletal maturation in
captive-born squirrel monkeys
(Saimiri boliviensis). Galliari, C. A. (Centro Argentino de
Primates, CONICET, Casilla de Correo 145, (3400) Corrientes, Argentina).
American Journal of Primatology, 1988, 16, 51-61.
. . This study is based on a mixed longitudinal radiographic sampling
of appendicular bones in 82 captive-born squirrel monkeys. All
appendicular ossification centers had appeared radiographically by 17
weeks of age, and epiphyseal fusion was complete by 47-53 months of age.
No statistically significant differences were found between the sexes in
comparisons carried out at birth and at 6 and 8 months of age.
*Elevation in plasma glucagon levels in response to stress in
squirrel monkeys: Comparisons of two subspecies
(Saimiri sciureus boliviensis and Saimiri sciureus sciureus) .
Meyers, B. A., Mendoza, S. P., & Cornelius, C. E. (S. P. Mendoza,
California Primate Research Center, Univ. of California, Davis, CA
95616). Journal of Medical Primatology, 1988, 17, 205-214.
. . Bolivian and Brazilian squirrel monkeys were captured and
restrained to determine patterns of glucagon and glucose response. In
both subspecies, plasma glucagon and glucose levels rose rapidly in
response to restraint procedures; glucagon levels in
boliviensis and glucose levels in both subspecies returned to basal
values in 15 minutes. The magnitude of the glucagon and glucose
responses differed significantly between the subspecies.
*Variation in rates of activity and inhibition of serum
cholinesterase among anthropoid primates. Breed, R. T. & Jolly, C. J.
(C. J. Jolly, Dept. of Anthropology, New York Univ., New York, NY
10003).
International Journal of Primatology, 1988, 9, 97-112.
. . Cholinesterase activity in the plasma of 5 hominoid and 5
cercopitheciod species was measured and characterized by profiles of
inhibition by dibucaine and NaF. Chimpanzees and gorillas strongly
resemble humans, and differ from other hominoids and cercopithecoids, in
patterns of dibucane and fluoride resistance and relative activity
toward various thio esters. "Silent" cholinesterase phenotypes of
probable veterinary significance, and analogous to a rare human variant,
are apparently normal in some catarrhine species and are present as a
polymorphism among orangutans and lion-tailed macaques.
*Endogenous rhythms in the succession of sleep-wake stages in
Microcebus murinus. Petter-Rousseaux, A. & Barre, V. (CNRS, Lab.
d'Anatomie Comparée, 55 rue Buffon, 75005 Paris, France).
International Journal of Primatology, 1988, 9, 147-163.
. . Sleep-wake stages were studied by means of EEG recordings in 3
female
Microcebus maintained under dim red light for 2 to 12 months and in
a female maintained in constant bright light first for 1 month, then for
4 months. A circadian rhythm was apparent in all cases. In addition, the
reduction of the alert-wake state in winter and its large increase in
summer hints at a circannual rhythm.
*Prenatal growth in the cynomolgus and rhesus macaque
(Macaca fascicularis and Macaca mulatta) : A comparison by
ultrasonography. Tarantal, A. F. & Hendrickx, A. G. (California Primate
Research Center, Univ. of California, Davis, CA 95616).
American Journal of Primatology, 1988, 15, 309-323.
. . Observations during the embryonic and fetal periods in cynomolgus
and rhesus macaques have been compared with diagnostic ultrasound.
Results indicate no significant differences in size during the embryonic
and early fetal periods, but a greater acceleration of growth in the
rhesus begins at approximately 100-110 gestational days. Analysis of
embryonic and fetal heart rates indicate no differences between the two
species. Normal predictive values for a variety of growth parameters
including gestational sac, greatest length, biparietal diameter, and
femur length have been calculated by multiple regression analysis. These
charts have proven useful for confirming the gestational age after timed
matings and for predicting the age of animals for which the conception
date is not known.
Reproduction
*International Cotton-top Tamarin Studbook. 1st Edition. S. D. Tardif & R. Colley. Oak Ridge: Oak Ridge Associated Universities, 1987. [Order from Marmoset Research Center, Oak Ridge Associated Universities, Oak Ridge, TN 37831]
*International Cotton-top Tamarin Studbook. 2nd Edition. S. D. Tardif & R. Colley. Oak Ridge: Oak Ridge Associated Universities, 1988. [Ordering information same as above]
*1987 Black Howler Monkey Regional Studbook. Barbara Baker. Columbia, SC: Riverbanks Zoological Park, 1988. 56pp.
*A preliminary study of olfactory behavior of captive
Lemur coronatus during the breeding season. Kappler, P. M. (Dept.
of Zoology & Primate Center, Duke Univ., Durham, NC 27705).
International Journal of Primatology, 1988, 9, 135-146.
. . The olfactory behavior of a captive population of crowned lemurs
was studied during their annual breeding season in order to investigate
its possible role in the fine-tuning of mating synchrony after
photoperiodic initiation of reproductive activity.
*Twinning among
Callimico goeldii. Altmann, J., Warneke, M., & Ramer, J. (Allee
Lab. of Animal Behavior, Univ. of Chicago, 940 E. 57th St., Chicago, IL
60637).
International Journal of Primatology, 1988, 9, 165-168.
. . One of the main features that distinguishes callimicos from
callitrichids is the production of only a single infant per gestation,
rather than the litter size of two that sets the Callitrichidae apart
from all other anthropoid primates. This is a report on 4 sets of twins
among 188 pregnancies among callimicos at Brookfield Zoo. In the one
pair that survived, one infant had to be hand-raised, while only one
infant in one other pair survived.
*Proximate and ultimate causes of reproductive suppression among
female yellow baboons at Mikumi National Park, Tanzania. Wasser, S. K. &
Starling, A. K. (Dept. of Psychology, Univ. of Washington, Seattle, WA
98195).
American Journal of Primatology, 1988, 16, 97-121.
. . Attack coalitions, whereby two or more individuals attack another
animal simultaneously, were studied for 14 months in 53 sexually mature
female baboons from one small and two large troops. Results suggest that
attack coalition behavior is a form of reproductive competition whereby
females attempt to suppress the reproduction of others at predictably
competitive times and thereby reduce the competition their own infants
face from the time they are weaned.
*Ejaculate quality, testes size and sperm competition in primates.
Møller, A. P. (Dept. of Zoology, Uppsala Univ., Box 561, S-751 22
Uppsala, Sweden).
Journal of Human Evolution, 1988, 17, 479-488.
. . Literature data on 25 primate species were examined for
correlations between measures of ejaculate quality and testes size and
body size, respectively. Ejaculate volume and sperm concentration were
positively correlated with body size in an interspecific analysis.
Testes size, after controlling for the effect of body size, was related
to breeding system in primates; species in which more than one male
copulated with each oestrous female had larger testes than those with
single-male breeding systems. Primate species with multi-male breeding
systems produced ejaculates with relatively higher sperm motility and
with relatively more motile sperm than did primates with single-male
systems.
*Coalitions, cooperation and reproductive tactics among adult male
baboons. Bercovitch, F. B. (Wisconsin Regional Primate Research Center,
Univ. of Wisconsin, 1223 Capitol Court, Madison, WI 53715).
Animal Behaviour, 1988, 36, 1198-1209.
. . Male savanna baboons,
Papio cynocephalus, form coalitions with each other as a technique
for gaining access to a female in consort with another male. Reciprocal
altruism has been invoked as the primary mechanism underlying this
reproductive tactic, but a 19-month field study of the reproductive
behavior of olive baboons,
P. c. anubis, in Kenya contradicted this interpretation. Males who
solicited others for coalition formation were not more likely to gain
access to consort females than were their fellow coalition members.
Males who refused to participate in a coalition at one time were still
sought as coalition partners at other times by the same male. Coalitions
were generally formed by older, middle- to lower-ranking males and
targeted at younger, higher-ranking males. Coalitions are a low risk
reproductive tactic and the benefits are not partitioned according to
initiator/joiner status.
*Alterations in primate sperm motility with maturation and during
exposure to theophylline. Gould, K. G., Young, L. G., & Hinton, B. T.
(Yerkes Regional Primate Research Center, Emory Univ., Atlanta, GA
30322).
American Journal of Primatology, 1988, 15, 325-336.
. . Data show that sperm from the caput epididymis of the chimpanzee
have the capacity for normal motility, but do not have the capacity to
bind to and penetrate an ovum, while sperm from its cauda epididymis
have the capacity for normal motility as well as the capacity to bind to
and penetrate an ovum.
*Puberty in primates. Plant, T. M. (Dept. of Physiology, Univ. of
Pittsburgh School of Medicine, Pittsburgh, PA 15261). In
E. Knobil, J. Neill et al. (Eds.).
The Physiology of Reproduction (pp. 1763-1788).
New York: Raven Press, 1988.
. . The study of puberty in primates has been restricted almost
exclusively to the infraorder, Catarrhini. In such higher primates,
puberty may be viewed as manifesting itself as a result of the cascade
of physiological and behavioral processes that are set in motion by the
reawakening of a dormant, but apparently fully mature, hypothalamic GnRH
pulse generator. Although physiological models with a central neural
timekeeping system or growth-tracking device have been proposed to
account for the timing of puberty, the mechanisms that dictate the
arrest and reawakening of the hypothalamic GnRH pulse generator during
postnatal life remain a fundamental problem in developmental
neurobiology. Studies of New World primates and prosimians are scant,
and therefore it is not presently possible to speculate whether the
timing of puberty in the lower primates is subserved by the same
mechanisms that govern this process in Old World monkeys, apes, and
humans.
*Progesterone levels during pregnancy in the greater thick-tailed
Galago
(Galago crassicaudatus) . Izard, M. K. & Fail, P. A. (Duke Univ.
Primate Center, Durham, NC 27705).
Journal of Medical Primatology, 1988, 17, 125-133.
. . Serum progesterone concentrations in
Galago crassicaudatus were measured at 3-week intervals throughout
the 136-day pregnancy. Progesterone concentrations were significantly
elevated over those of nonpregnant controls as early as 6 weeks after
conception, and continued to increase throughout gestation. The
progesterone profile in pregnant
G. crassicaudatus quantitatively resembles that of chimpanzees and
qualitatively resembles that of humans. In two animals that aborted,
progesterone concentrations after abortion decreased to values
comparable to those seen in nonpregnant animals.
*The primate oviduct and endometrium. Brenner, R. M. & Maslar, I. A.
(Oregon Regional Primate Center, 505 NW 185th Ave, Beaverton, OR 97005).
In E. Knobil, J. Neill et al. (Eds.),
The Physiology of Reproduction (pp. 303-329). New York: Raven, 1988.
. . The cyclic fluctuations in serum estradiol and progesterone that
occur during the primate menstrual cycle are clearly responsible for the
dramatic, cyclic changes that occur throughout the reproductive tract.
This review focuses on the similarities and differences in hormonal
responsiveness that exist between the primate endometrium and oviduct.
*Infant mortality in mother-reared captive chimpanzees at Taronga
Zoo, Sydney. Courtenay, J. (Dept. of Zoology, Australian National Univ.,
Canberra City, ACT, 2601, Australia).
Zoo Biology, 1988, 7, 61-68.
. . Sex-specific mortality probabilities were calculated for
mother-reared chimpanzees using standard demographic techniques with
Taronga Zoo records. Male mortality was found to be considerably higher
than female mortality. Changes in infant mortality over time were
calculated, with particular reference to changes that had occurred since
the chimpanzees were transferred to a new open air enclosure in 1980.
Mortality was found to have decreased dramatically since the move to the
new enclosure, but the observed decline was exaggerated by the very high
proportion of female infants. The main causes of infant death were
pneumonia and trauma. In addition, undetermined causes in the first day
of life were numerous. An examination of the contexts of infanticide in
wild and captive chimpanzees revealed slight differences, but factors
such as aggression towards unfamiliar individuals and redirected
aggression were common to both. Cannibalism in the zoo has so far been
limited to the bodies of stillborn infants.
*Responses of foster-mothers and troop members to adopted newborns
in a captive group of rhesus monkeys. Holman, S. D. & Goy, R. W. (MRC
Neuroendocrine Development and Behaviour Group, AFRC Inst. of Animal
Physiology, Babraham, Cambridge CB2 4AT, England).
Primates, 1988, 29, 343-352.
. . Two male and two female rhesus monkeys between 24 and 120 hours
old, born to females caged singly, were exchanged with same-sex infants
born to females in a large troop which was held in an enclosure in a
different building. Both foster-mothers and other troop members
recognized the unfamiliarity and novelty of the foster infant, but the
infants were still accepted into the group.
*Social correlates of reproductive suppression in captive
callitrichid family groups. Carroll, J. B. (Jersey Wildlife Preservation
Trust, Les Augres Manor, Trinity, Jersey, Channel Islands).
Dodo, 1986, 23, 80-85.
. . Case histories from the records of the callitrichid colony at
Jersey Wildlife Preservation Trust, which suggest that familial
relationship between father and daughter or mother and son is important
in suppression of breeding, rather than dominance relationships between
mother and daughter or father and son.
Taxonomy
*Gentle lemurs: New species, and how they are formed. Groves, C. P.
(Dept. of Prehistory & Anthropology, Australian National Univ.,
Canberra, ACT 2601, Australia).
Australian Primatology, 1988, 3[2/3], 9-12.
. . Descriptions of, and comments on, the characterization, taxonomy,
evolution, and conservation, of the species and subspecies of
Hapalemur.
*Systematics of the Lemuridae (Primates, Strepsirhini). Groves, C.
P. & Eaglen, R. H. (Dept. of Prehistory & Anthropology, Australian
National Univ., Canberra ACT 2601, Australia).
Journal of Human Evolution, 1988, 17, 513-538.
. . After reviewing the literature and examining museum specimens
of the family Lemuridae, the authors suggest the elevation of the
fulvus group to the rank of genus, and propose the name Petterus
for it, with type P. fulvus. The incorporation of Petterus
in the family Lemuridae, without including any suprageneric groupings,
provides a way of indicating the present uncertainty about the
phylogenetic affinities among its members.
*Molecular systematics of higher primates: Genealogical relations
and classification. Miyamoto, M. M., Koop, B. F., Slightom, J. L.,
Goodman, M., & Tennant, M. R. (Dept. of Zoology, Univ. of Florida,
Gainesville, FL 32611).
Proceedings of the National Academy of Science, 1988, 85,
7627-7631.
. . The authors obtained 5' and 3' flanking sequences (5.4 kilobase
pairs) from the psi-eta-globin gene region of
Macaca mulatta and combined them with
available nucleotide data. The completed sequence, representing 10.8
kilobase pairs of contiguous noncoding DNA, was compared to the same
orthologous regions available for human
(Homo sapiens, as represented by 5 different alleles), common
chimpanzee (Pan troglodytes), gorilla (Gorilla gorilla), and
orangutan (Pongo pygmaeus). The nucleotide sequence for Macaca
mulatta provided the outgroup perspective needed to evaluate better the
relationships of humans and great apes. Pairwise comparisons and
parsimony analysis of these orthologues clearly demonstrated that humans
and great apes share a high degree of genetic similarity and that
humans, chimpanzees, and gorillas form a natural monophyletic group.
These conclusions strongly favor a genealogical classification for
higher primates consisting of a single family (Hominidae) with two
subfamilies (Homininae for Homo, Pan, and Gorilla,
and Poninae for Pongo).
Taxonomic status of
Saguinus tripartitus (Milne-Edwards, 1878). Thorington, R. W. Jr.
(NHB 390, Smithsonian Institution, Washington, DC 20560).
American Journal of Primatology, 1988, 15, 367-371.
. . A review of specimens and field notes show that
Saguinus tripartitus and S. fuscicollis lagonotus are sympatric,
although they probably occur in different habitats. Therefore,
S. tripartitus should be treated as a distinct species, not as a
subspecies of
S. fuscicollis.
-------------------------------------------------------------------
In many cases, the original source of references in this section has been the Current Primate References prepared by The Primate Information Center, Regional Primate Research Center SJ-50, University of Washington, Seattle, WA 98l95. Because of this excellent source of references, the present section is devoted primarily to presentation of abstracts of articles of practical or of general interest. In most cases, abstracts are those of the authors.
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. (Phone: 401-863-2511)
Judith_Schrier@brown.edu
ACKNOWLEDGMENTS
The Newsletter is supported by U. S. Public Health
Service Grant RR-00419 from the Animal Resources Program,
Division of Research Resources, N.I.H.
Cover drawing of an infant marmoset (Callithrix jacchus) by Penny Lapham.
Copyright © 1989 by Brown University
Editor: Judith E. Schrier, M. Sc.
Associate Editor: James S. Harper, D.V.M.
Consulting Editor: Morris L. Povar, D.V.M.
Founding Editor: Allan M. Schrier, Ph.D.
{kind=link}
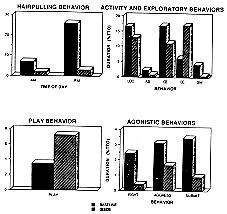{kind=link}
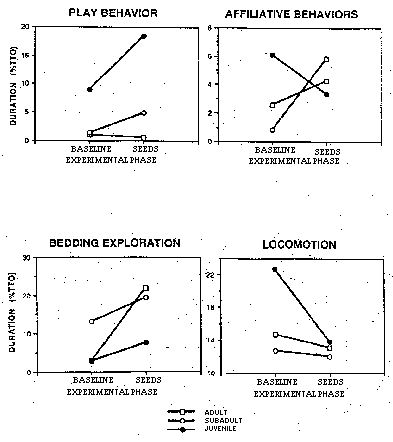{kind=link}
{kind=link}
{kind=link}